- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Colocalization Analysis for Cryosectioned and Immunostained Tissue Samples with or without Label Retention Expansion Microscopy (LR-ExM) by JACoP
Published: Vol 12, Iss 5, Mar 5, 2022 DOI: 10.21769/BioProtoc.4336 Views: 4784
Reviewed by: Nafisa M. JadavjiHai HuangKai Yuan

Original research article
The authors used this protocol in:

Jun 2021
Advertisement


Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics







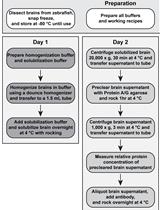
Abstract
Asymmetric cell division (ACD) is fundamental for balancing cell proliferation and differentiation in metazoans. During active neurogenesis in the developing zebrafish forebrain, radial glia progenitors (RGPs) mainly undergo ACD to produce one daughter with high activity of Delta/Notch signaling (proliferative cell fate) and another daughter with low Delta/Notch signaling (differentiative cell fate). The cell polarity protein partitioning-defective 3 (Par-3) is critical for regulating this process. To understand how polarized Par-3 on the cell cortex can lead to differential Notch activity in the nuclei of daughter cells, we combined an anti-Delta D (Dld) -atto 647N antibody uptake assay with label retention expansion microscopy (LR-ExM), to obtain high resolution immunofluorescent images of Par-3, dynein light intermediate chain 1 (Dlic1), and Dld endosomes in mitotic RGPs. We then developed a protocol for analyzing the colocalization of Par-3, Dlic1, and endosomal DeltaD, using JACoP (Just Another Co-localization Plugin) in ImageJ software (Bolte and Cordelières, 2006). Through such analyses, we have shown that cytosolic Par-3 is associated with Dlic1 on Dld endosomes. Our work demonstrates a direct involvement of Par-3 in dynein-mediated polarized transport of Notch signaling endosomes. This bio-protocol may be generalizable for analysis of protein co-localization in any cryosectioned and immunostained tissue samples.
Keywords: Par-3Background
Asymmetric cell division (ACD) of progenitor/stem cells is critical for generating cellular diversity in tissue differentiation and organogenesis during development (Chenn and McConnell, 1995; Fishell and Kriegstein, 2003; Williams et al., 2011). During ACD, each daughter cell acquires distinct fates after mitosis by asymmetric inheritance of cellular components (so-called cell fate determinants), e.g., one continuing to self-renew and the other embarking on a differentiation path (Horvitz and Herskowitz, 1992; Knoblich, 2001). Previous studies from our laboratory and others have shown that cell polarity protein Par-3 is required to establish the asymmetric activity of Notch signaling between daughter cells (Notchhi vs. Notchlo) (Mizutani et al., 2007; Bultje et al., 2009; Coumailleau et al., 2009; Dong et al., 2012; Derivery et al., 2015; Kressmann et al., 2015), a key regulator of cell fate decisions (Artavanis-Tsakonas et al., 1999; Gaiano and Fishell, 2002). Recent studies show that Notch signaling in stem cell division is heavily controlled by endocytic trafficking (Chitnis, 2006; Daeden and Gonzalez-Gaitan, 2018). In Drosophila sensor organ precursor (SOP) cells, Notch signaling endosomes undergo directional movement (Derivery et al., 2015). Built on these previous findings, we investigated how Par-3 might regulate the dynamics of Dld endosomes in mitotic RGPs (Zhao et al., 2021).
In this protocol, we describe our method for visualizing and analyzing cytoplasmic Par-3 in association with the dynein complex subunit Dlic1 and Dld endosomes in zebrafish RGP cells. The antibody uptake assay was used to label Dld endosomes in zebrafish embryos (Tong et al., 2019; Zhao et al., 2021). Following live labeling, cryosectioning, immunofluorescent staining, and label retention microscopy (LR-ExM) (Shi et al., 2021) were carried out. We have optimized these procedures and the protocol detailed here should be generalizable to all cryosectioned and immunostained tissue samples, not limited to zebrafish. Subsequently, we obtained high-resolution images of mitotic RGP cells in the processed samples, using spin-disk confocal microscopy. The cytoplasmic areas between two nuclei of the anaphase/telophase RGPs were designated as the regions of interest (ROIs) for colocalization analyses of Par-3, Dlic1, and Dld. Because the size of endosomes (average diameters of 250 nm to 1000 nm) is much larger than the pixel size of fluorescent images, the fluorescence signals colocalized on Dld endosomes may not appear to exactly overlap with one another. Based on previously published reports (Bolte and Cordelières, 2006; Derivery et al., 2015), we developed the method for measuring co-localization in the context of endosomes in this protocol. For all three channels, we measured the colocalization between every two fluorescent channels in the form of Manders’ Coefficients (i.e., the proportion of each fluorescence colocalized with another) by using the JACoP plugin in ImageJ (Manders et al., 1993). Our results uncovered that Par-3 is colocalized with Dlic1 on Dld endosomes, hence revealing a direct role of cytoplasmic Par-3 in actively localizing intracellular determinants during ACD of RGP cells (Zhao et al., 2021).
Materials and Reagents
FisherbrandTM SuperfrostTM Plus microscope slides (Fisher Scientific, catalog number: 22-037-246)
6-well glass bottom plates (CellVis, catalog number: P06-1.5H-N)
Zebrafish strains wild-type zebrafish embryos were obtained from natural spawning of AB adults, staged and maintained according to established protocols (Kimmel et al., 1995)
Zebrafish embryos were raised at 28.5 °C in 0.3× Danieau’s embryo medium (Zhao et al., 2021)
Capillaries, 1.2 mm OD, 0.9 mm ID, with filament (World Precision Instruments, catalog number: 1B120F-6)
Tricaine (Sigma-Aldrich, CAS Number: 886-86-2, catalog number: E10521)
Phenol red (Sigma-Aldrich, catalog number: P0290)
Primary antibodies used in this study:
Mouse anti-Dld (Abcam, catalog number: ab73331; RRID, catalog number: AB_1268496; lot, catalog number: GR115501-3, 1:200 dilution for immunostaining)
Chicken anti-GFP (Abcam, catalog number: ab13970; RRID:AB_300798, lot GR3190550-20, 1:500 dilution for immunostaining)
Rabbit anti-Par-3 (Millipore 07-330; RRID:AB_2101325; lot 3322358, 1:500 for immunostaining)(validated in this study)
Guinea pig anti-DLIC1-Cter (a gift from Dr. T. Uemura, 1:100 for immunostaining) (Satoh et al., 2008)
Secondary antibodies used for immunofluorescent labeling:
Alexa®-conjugated goat anti-rabbit (Alexa 568, Invitrogen, catalog number: A11011, RRID. AB_143157, lot 792518, 1:2,000 dilution)
Goat anti-chicken (Alexa 488, Invitrogen, catalog number: A11039, RRID:AB_142924, lot 2020124, 1:2,000 dilution)
Goat anti-mouse (Alexa 488, Invitrogen, catalog number: A11002, RRID:AB_2534070, lot 1786359, 1:2,000 dilution)
Goat anti-guinea pig (Alexa 488, Invitrogen, catalog number: A11073, RRID:AB_2534117, lot 46214A, 1:2,000 dilution) or Donkey anti-guinea pig (Alexa 647, Jackson Labs, catalog number: 706-605-148, RRID:AB_2340476, lot 102649-478, 1:2,000 dilution)
Anti-Mouse-IgG-Atto647N (Sigma-Aldrich, catalog number: 50185, 1 mg/mL)
Secondary antibodies used for tri-functional linker conjugation in LR-ExM were:
Goat anti-Guinea Pig IgG (H+L) unconjugated secondary antibody (Invitrogen, catalog number: A18771, RRID:AB_2535548)
Goat anti-Rabbit IgG (H+L) Cross-Adsorbed unconjugated secondary antibody (Invitrogen, catalog number: 31212, RRID:AB_228335)
Goat anti-Chicken IgY (H+L) unconjugated secondary antibody (Invitrogen, catalog number: A16056, RRID:AB_2534729)
NHS-MA-Biotin conjugated anti-Chicken IgY, NHS-MA-Biotin conjugated anti-Rabbit IgG, and NHS-MA-DIG conjugated anti-Guinea Pig IgG (Gift from Dr. Shi) (Shi et al., 2021)
DAPI solution (1 mg/mL) (Thermo ScientificTM, catalog number: 62248)
Tween 20 (Sigma-Aldrich, CAS Number: 9005-64-5, catalog number: P-9416)
TritonTM X-100 (Sigma-Aldrich, CAS Number: 9036-19-5)
Dimethyl sulfoxide (DMSO) (Sigma-Aldrich, CAS Number: 67-68-5)
Natural goat serum (Invitrogen, catalog number: 31872)
Glycerol (Sigma-Aldrich, CAS Number: 56-81-5G, catalog number: 9012)
Phosphate-buffered saline (PBS) 10× solution, pH 7.4 (GibcoTM, catalog number: 70011044)
Sucrose (Sigma-Aldrich, CAS number: 57-50-1)
Tissue-PlusTM O.C.T. compound (Fisher Scientific, catalog number: 23-730-571)
UltraPureTM low melting point agarose (Invitrogen, catalog number: 16520050)
Sodium acrylate (Sigma-Aldrich, CAS Number: 7446-81-3)
Acrylamide (Sigma-Aldrich, CAS Number: 79-06-1, catalog number: A9099)
Sodium chloride (Sigma-Aldrich, CAS Number: 7647-14-5, catalog number: 204439)
N,N,N′,N′-Tetramethyl ethylenediamine, TEMED (Sigma-Aldrich, CAS Number: 110-18-9, catalog number: 1.10732)
Ammonium persulfate, APS (Thermo ScientificTM, catalog number: 17874)
Proteinase K (Sigma-Aldrich, CAS Number: 39450-01-6, catalog number: P4850)
N,N′-Methylenebis-acrylamide (Sigma-Aldrich, CAS Number: 110-26-9, catalog number: M7279)
30× Danieau’s embryo medium (see Recipes)
Blocking buffer for immunostaining (see Recipes)
Washing buffer (see Recipes)
LR-ExM gelation buffer and digestion buffer (see Recipes)
Gelation buffer (see Recipes)
Monomer solution (see Recipes)
Digestion buffer (see Recipes)
Post-digestion staining buffer (see Recipes)
Equipment
Stero dissection microscope (Zeiss Stemi 2000 with a maximum magnification of 50)
Air pressure injector( Narishige IM 300 microinjector)
Micromanipulators (WPI M3301R, World Precision Instruments, Sarasota, FL, USA)
Flaming-Brown P897 needle puller (Sutter Instruments, Novato, CA, USA)
Nikon CSU-W1 Spinning Disk/confocal microscope
Leica CM1850 Cryostat
Software
Micro-Manager 2.0 gamma (μManagerTM, University of California), https://micro-manager.org/
Fiji (Schindelin et al., 2012), https://imagej.net/software/fiji/
Prism 9, GraphPad, https://www.graphpad.com/
Procedure
Anti-Dld antibody uptake assay in the one day post fertilization (1dpf) zebrafish brain
Antibody conjugation: Mix 1 μL of anti-Dld antibody (0.5 mg/mL) with 2.5 μL of Anti-Mouse-IgG-Atto647N antibody (1 mg/mL) and incubate at room temperature for 30 min, or on ice for 2–3 h. From this step, the mixture should be protected from light.
Blocking of unconjugated antibody: Add 2.5 μL of blocking buffer (10 mg/mL of mouse IgG with 5 mM azide, supplied by Sigma-Aldrich with the order of anti-Mouse-Atto647N) and 0.5 μL of 0.5% Phenol red (Sigma P0290) for blocking the unconjugated Anti-Mouse-IgG-Atto647N. Vortex thoroughly, and incubate the mixture at room temperature for 5–10 min. Mixtures without anti-Dld antibody were used as control.
Microinjection: Anaesthetize 24–26 h post fertilization (hpf) embryos in the Danieau medium supplemented with 0.003% tricaine, followed by embedding in 1.2% low-melt point agarose. The zebrafish microinjection platform is the same as previously described (Dong et al., 2011). Inject 10 nL of labelled Dld antibody into the hindbrain ventricle. Release the injected embryos from agarose and culture them in the Danieau medium for 3–4 h.
Before proceeding to fixation and immunostaining, monitor the effects of the antibody uptake assay under a spinning-disk confocal microscope.
Fixation and cryosectioning of zebrafish embryos
Collect embryos from the last step, wash them in PBS twice, and then transfer them to Eppendorf tubes with 4% paraformaldehyde (PFA) overnight at 4°C in 1× PBS buffer.
Wash the fixed embryos three times with 1× PBS (5 min each time) and incubate them in 1× PBS buffer containing 30% sucrose overnight at 4°C.
Transfer the embryos to plastic molds for preparing frozen samples with soft tips, remove the sucrose buffer, and add the OCT (Tissue-tek). After orienting the embryos to proper positions in the mold, freeze the block on dry ice. Blocks can be stored at -80°C up to several months.
Cut frozen blocks into 20 μm sections on a cryostat (Leica). After placing the sections on Superfrost Plus slides (Fisher Scientific), these slides were dried at room temperature for 2–3 h and then stored at -80°C until use.
Whole mount fluorescent immunostaining
Wash the frozen cryosections in PBS-T for 5 min, then preincubate them in blocking buffer at 4°C overnight or longer.
After blocking, incubate the cryosections with primary antibodies in the blocking buffer overnight at 4°C.
For those samples that do not continue with LR-ExM, thoroughly wash the cryosections with PBS-T five times × 10 min each time, and incubate with Alexa®-conjugated goat anti-rabbit (Alexa 568), goat anti-chicken (Alexa 488), goat anti-mouse (Alexa 488), or goat anti-guinea pig (Alexa 647) secondary antibodies (diluted 1:1,000) in the blocking buffer, for over 2 h at room temperature or overnight at 4°C.
Wash the samples with PBS-T twice, and then three times with PBS (10 min each).
Incubate the samples with 50% glycerol in PBS for 1 h, followed by infiltration overnight in 80% glycerol/PBS before imaging.
Label Retention Expansion Microscopy (LR-ExM)
For the cryosections to be used for LR-ExM, after primary antibody incubation as described above, they were incubated with trifunctional linkers [NHS-MA-Biotin conjugated anti-Chicken (or anti-Rabbit) IgG—for visualizing Par-3, and NHS-MA-DIG conjugated anti-Guinea Pig IgG—for visualizing Dlic1 (200 mg/mL stock, dilute 1:100 before use)] in the blocking buffer overnight at 4°C in the dark.
After washes in PBS (5 min each) in the dark, add 40 μL of freshly prepared gelation solution onto each section to cover the whole tissue sample. The gelation solution was prepared by deoxygenizing the gel monomer solution using a vacuum pump for over 15 min, before adding TEMED and APS, to enhance the effects of trifunctional linkers.
Protect the sections from light, incubate in a humidity chamber, and allowe to undergo gelation at 37°C for 1 h.
Incubate the gelated samples in the digestion buffer (see Recipes) on the slides at 37°C for 4 h or at room temperature overnight. At least 10-fold excess volume of digestion buffer was used (> 5 mL for each slide). Sufficient digestion enables sections embedded in gels to slide off the glass surface.
Wash the gel embedded samples with excess volume of 150 mM NaCl in 6-well plates of at least 5 mL in each well for four times (20–30 min each time).
After washing off the digestion buffer, incubate the gel samples in the post-digestion staining buffer (see Recipes) with 3–5 μM Alexa Fluor® 488-Streptavidin, 3–5 μM goat anti-Digoxigenin/Digoxin Dylight® 594, anti-mouse Atto647N (1:500), and DAPI (1:1,000) in the dark at 4°C for 24 h.
Wash the gel embedded samples four times with Milli-Q water (30 min for each wash) in the dark at 4°C. The samples expand to approximately four times their original size after washing and are then ready for imaging.
Confocal microscopy
Immuno-stained samples without LR-ExM were scanned using a confocal microscope (Nikon CSU-W1 Spinning Disk/High Speed Widefield confocal microscopy, Nikon Imaging Core at UCSF) at 0.26 μm z-step size, with a 100× Oil objective. LR-ExM samples were scanned with the same confocal microscope at 0.26 μm z-step size, with a 60× water immersion (WI) objective.
Data analysis
The colocalization of cytosolic Par-3 with Dlic1 and Dld endosomes in mitotic RGP was described in our recent publication (Zhao et al., 2021). We chose the central plane area between the nuclei of two nascent daughter cells as the Region of Interest (ROI) for colocalization analyses of Par-3, Dlic1, and Dld endosomes in the acquired images.
The acquired images of immuno-stained RGP cells in the zebrafish embryonic forebrain were segmented, and maximum intensity projections of ten consecutive z-planes covering the central cytoplasmic area of the cells were generated (0.26 μm z-step interval, the Max Z plane covered 2.6 μm thickness).
For the images of immuno-stained samples, the region of 60 × 100 pixels (1 pixel = 0.065 μm) between two nuclei of dividing RGP daughter cells was cropped and saved for colocalization analysis in FIJI (Fiji Is Just ImageJ) (Figure 1A).
For the images of LR-ExM processed samples, the area of 120 × 200 pixel (1 pixel = 0.11 μm) in the central cytoplasmic area between the nuclei was selected for JACoP colocalization analyses.
JACoP (Just Another Co-localization Plugin) was intalled into FIJI, if it was not in the list of Plugins. We then split the channel of the cropped images and chose those three channels for colocalization analyses to display on the desktop (Figure 1B).
JACoP can be found and downloaded from https://imagej.nih.gov/ij/plugins/track/jacop2.html
Running JACoP in FIJI for colocalization analysis
Open a cropped out image from step A.
If they are merged, split the channels in a compositive tiff file (Figure 1B).
Image>Color>Split Channels
Open JACoP
Plugins>JACoP (Figure 1C).
Click it to open a new window of JACoP plugin under FIJI (Figure 2).

Figure 1. Preparation of segmented RGP images for colocalization analyses. A. The central cytoplasmic area between the dividing cell nuclei was cropped and saved (the white rectangle in the image is 60 × 100 pixel. The size was shown by the red line highlighting w and h in FIJI). B. The image was split into three single channel images. C. Locating and choosing JACoP in the “Plugins” of FIJI.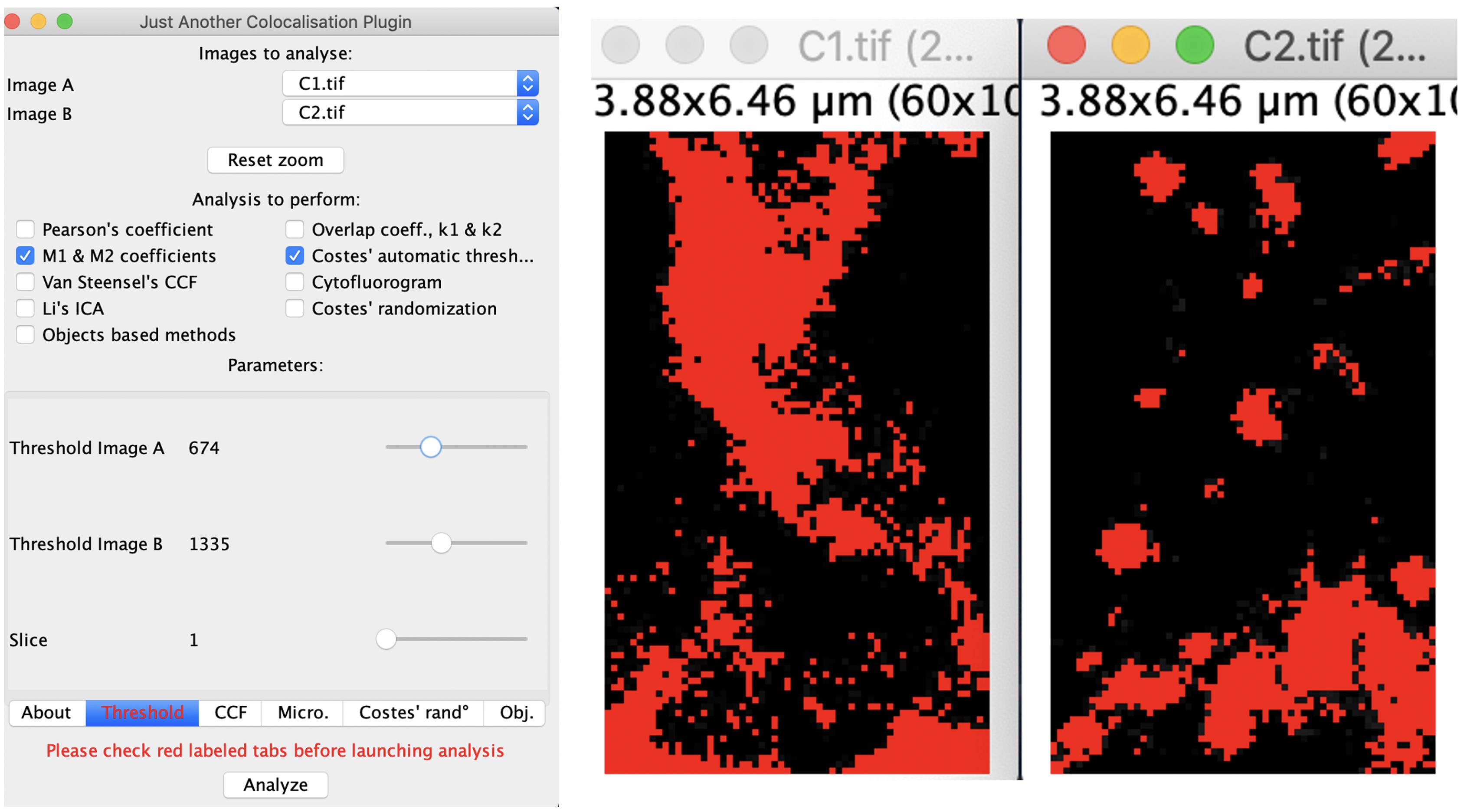
Figure 2. The interface for choosing channels, M1 & M2 coefficients, threshold, and Costes’ automatic threshold. The two images in C1 and C2 illustrate the effect after setting the thresholds.At the top of the window, select and compare the images of two different channels (C1 & C2) already opened by FIJI under “Image A” and “Image B” (left panel in Figure 2).
Then select the M1 & M2 coefficients (Manders’ coefficients) analysis to perform in the window of “Just Another Colocalisation Plugin” (left panel in Figure 2).
Check the tabs in Red for required settings before applying the analysis. To choose the correct functions in JACoP for analyses, please refer to the introduction of the plugin (Cordelieres and Bolte, 2008). In our study, we are only interested in Manders’ coefficients. CCF (cross-correlation coefficient) is not used for our study and would not be discussed here.
Threshold (at the bottom of left panel in Figure 2)
This is the threshold set for independent experiment. To remove potential background noise, the threshold of each channel was set in JACoP using a blank area on the image (i.e., without tissue samples) as a negative control.
You can determine the numbers consistently using the Image>Adjust>Threshold and use a predetermined automatic threshold, according to the masking area shown in each channel (right panel of Figure 2).
Costes’ automatic threshold was normally applied for each measurement, which is an algorithm to identify and remove noise using scatter plotting of randomized images generated from the image under analysis (Costes et al., 2004). It would generate another pair of automatically quantified M1 & M2 coefficients.
For negative control images, the M1 & M2 coefficients would be “0” using both thresholds.
Micro.
This is the information about your microscope (Figure 3A).
If you are using the image acquired from the Bio-imaging software (an OME.TIF or a .nd2, etc.), that information should be automatically associated with your image (xy calib nm, 65; z calib nm, 260, for the 100× objective).
Enter in Wavelength excitation information.
Costes’ rand°
The minimum “xy block size” can be set at 2. “z block size” was always set at 1, as we analyzed the Max-Z projection of image stacks (Figure 3B).
You can estimate this, for example:
1)All the pictures collected from the confocal microscope have the same size of 2048 × 2048 pixel, and the pixel size of the image is approximately 65 nm.
2)Typically, the size of endosomes is larger than 200 nm.
3)To cover the distance of 200 nm, we need at least 2 pixels from both channels.
4)When using images from LR-ExM samples, the minimum “xy block size” was usually set to 4. Because the sample was expanded for about 4 times, the minimum size of endosomes in the sample was enlarged to be over 800 nm. As we used a 60× WI objective for scanning LR-ExM samples, this increased the pixel size of the image to be about 110 nm. Therefore, to cover a distance of 800 nm, we need at least 4 pixels from both channels.
Obj. is the tab for checking parameter settings. No change should be required.
After going through each step, click “Analyze” at the bottom (Figure 3). The results are generated in a new window.
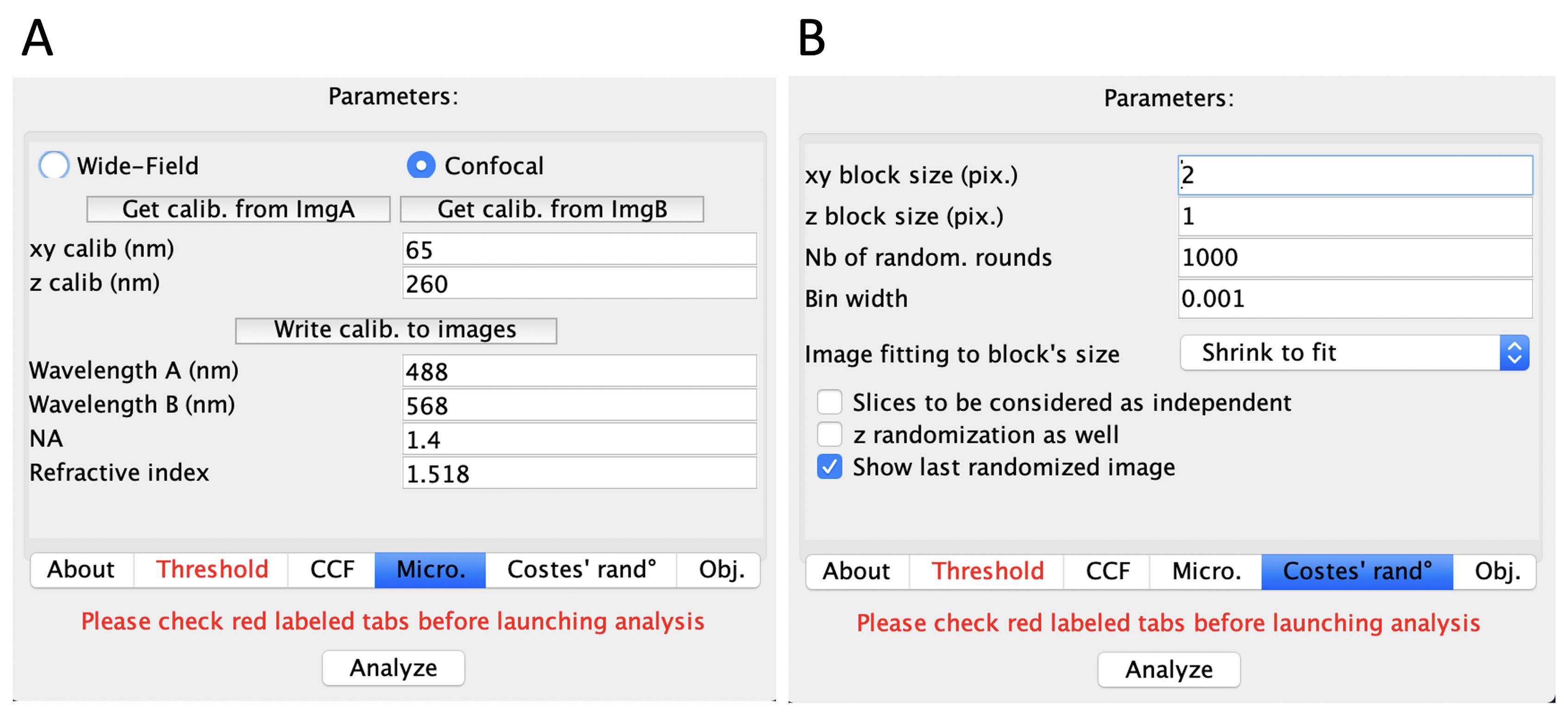
Figure 3. Demonstration of JACoP interface when choosing different tabs. A. Micro. displays the information of microscopy. B. Costes’ rand° displays the selection of xy block size for analysis.
To compare all three channels, the above steps should be repeated in pairs of two channels. All the results from the same sample are saved together in one txt. file. To achieve enough repeats for statistical analyses, the above steps were repeated for each image from all cells.
The statistics of Par-3, Dlic1, and Dld endosomes colocalization in each group of samples were assessed by Prism 9. For the images from the same experiment, M1 & M2 coefficients of Costes’ automatic threshold were also been used for comparison. For all images from independent experiments, we only used M1 & M2 coefficients of predetermined thresholds for comparison.
The colocalization relationship of those three proteins has been more clearly demonstrated by LR-ExM samples. As shown in Figure 4, the size of RGPs that have undergone LR-ExM is almost four times larger than normal immunostained RGPs shown in Figure 1. The colocalization analyses showed that similar colocalization coeffiecients for the three proteins have been found from both sets of data (Zhao et al., 2021).
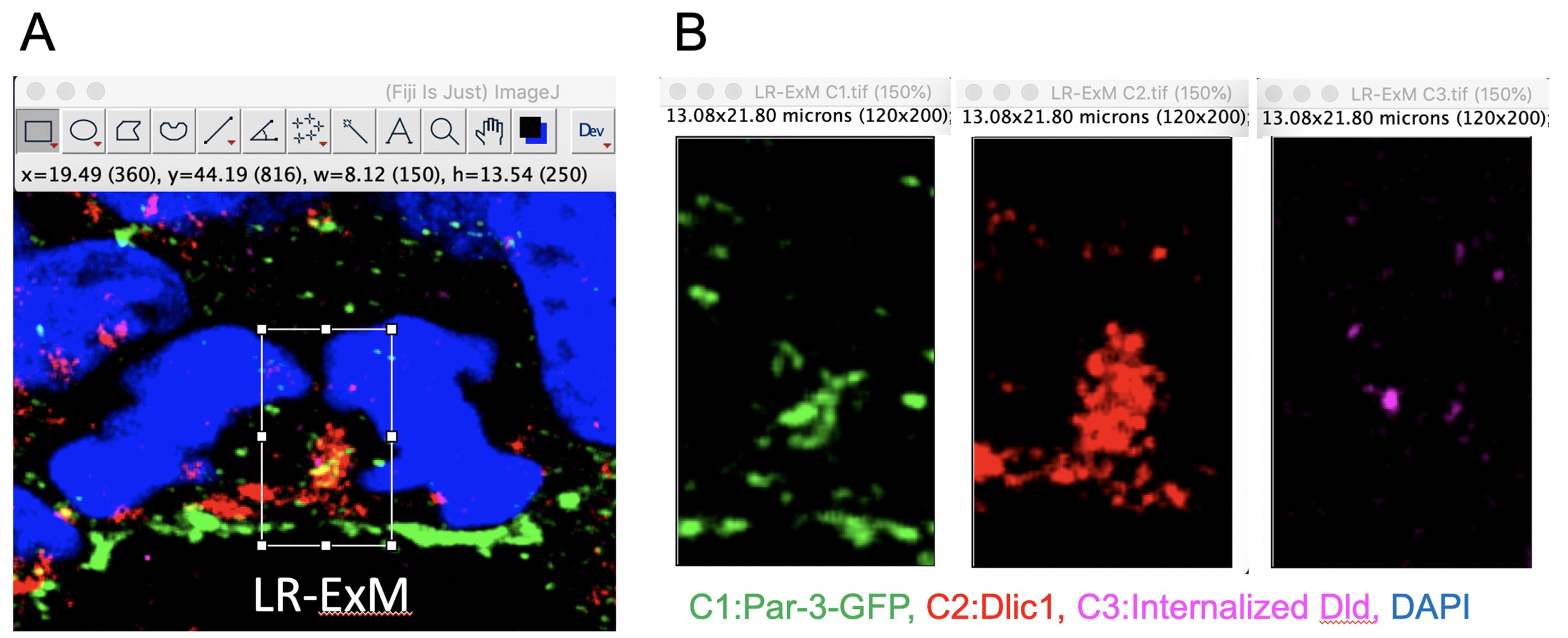
Figure 4. Preparation of segmented images of LR-ExM for colocalization analyses. A. The central cytoplasmic area between the dividing cell nuclei was cropped and saved (the white rectangle in the image is 150 × 250 pixel. The size was shown by w and h in FIJI. This picture is modified from (Zhao et al., 2021), some rights reserved; exclusive licensee AAAS. Distributed under a CC BY-NC 4.0 License (http://creativecommons.org/licenses/by-nc/4.0/). B. The image of the white rectangle area in A was split into three single channel images without showing DAPI. The size of the segmented area has been shown on the top of each frame.
Notes
The gelation buffer for LR-ExM should only be prepared freshly to achieve stable effects.
All the secondary antibodies conjugated with fluorophores were centrifuged and aliquoted (20 μL per tube) into light shielded Eppendorf tubes, and stored in a -80°C freezer.
Trifunction linkers for LR-ExM can only be stored at 4°C and shielded from light.
Recipes
30× Danieau’s embryo medium
1740 mM NaCl, 21 mM KCl, 12 mM MgSO4·7H2O, 18 mM Ca(NO3)2, 150 mM HEPES buffer
Blocking buffer for immunostaining
0.1% Tween 20, 0.25% Triton X-100 with 1% dimethyl sulfoxide (DMSO) and 5% normal goat serum in 1× PBS, pH 7.4.
Washing buffer
1× PBS with 0.05% Tween 20
Post-digestion staining buffer
10 mM HEPES, 150 mM NaCl in MilliQ water, pH 7.5
Monomer solution:
Sodium acrylate 0.86 g/mL, Acrylamide 0.25 g/mL, N,N′-Methylenebis-acrylamide 0.015 g/mL, Sodium chloride 1.17 g/mL in PBS (1×).
LR-ExM gelation buffer and digestion buffer are prepared by following the instruction in the original publication (Shi et al., 2021):
Gelation buffer
190 μL of Monomer solution with 5 μL of 8% TEMED and 5 μL 8% APS.
Digestion buffer
8 units/mL of Proteinase K in 50 mM Tris pH 8.0, 1 mM EDTA, 1 M NaCl, 0.5% of Triton X-100
Acknowledgments
This protocol was adapted from the previously published paper (Zhao et al., 2021). We thank DeLaine Larsen, Kari Herrington and UCSF Nikon imaging center for assistance with imaging and data analysis. We also thank Dr. Tadashi Uemura and Dr. Xiaoyu Shi for sharing the DLIC antibody and trifunctional linkers required for immunostaining and LR-ExM respectively. The project was supported by NIH R01NS120218 and the UCSF Mary Anne Koda-Kimble Seed Award for Innovation.
Competing interests
The authors declare no competing interests.
Ethics
All animal experiments were approved by the Institutional Animal Care and Use Committee (IACUC) at the University of California, San Francisco, USA.
References
- Artavanis-Tsakonas, S., Rand, M. D. and Lake, R. J. (1999). Notch signaling: cell fate control and signal integration in development. Science 284(5415): 770-776.
- Bultje, R. S., Castaneda-Castellanos, D. R., Jan, L. Y., Jan, Y. N., Kriegstein, A. R. and Shi, S. H. (2009). Mammalian Par3 regulates progenitor cell asymmetric division via notch signaling in the developing neocortex. Neuron 63(2): 189-202.
- Bolte, S. and Cordelières, F. P. (2006). A guided tour into subcellular colocalization analysis in light microscopy. J Microsc 224(3): 213-232.
- Chenn, A. and McConnell, S. K. (1995). Cleavage orientation and the asymmetric inheritance of Notch1 immunoreactivity in mammalian neurogenesis. Cell 82(4): 631-641.
- Chitnis, A., (2006). Why is delta endocytosis required for effective activation of notch. Dev Dyn 235(4): 886-894.
- Cordelieres, F. P. and Bolte, S. (2008). JACoP v2. 0: improving the user experience with co-localization studies. Proceedings of the 2nd ImageJ user and developer conference pp. 6-7.
- Costes, S. V., Daelemans, D., Cho, E. H., Dobbin, Z., Pavlakis, G. and & Lockett, S. (2004). Automatic and quantitative measurement of protein-protein colocalization in live cells. Biophys J 86(6): 3993-4003.
- Coumailleau, F., Furthauer, M., Knoblich, J. A. and Gonzalez-Gaitan, M. (2009). Directional Delta and Notch trafficking in Sara endosomes during asymmetric cell division. Nature 458(7241): 1051-1055.
- Daeden, A. and Gonzalez-Gaitan, M. (2018). Endosomal Trafficking During Mitosis and Notch-Dependent Asymmetric Division. Prog Mol Subcell Biol 57: 301-329.
- Derivery, E., Seum, C., Daeden, A., Loubery, S., Holtzer, L., Julicher, F. and Gonzalez-Gaitan, M. (2015). Polarized endosome dynamics by spindle asymmetry during asymmetric cell division. Nature 528(7581): 280-285.
- Dong, Z., Wagle, M. and Guo, S. (2011). Time-lapse live imaging of clonally related neural progenitor cells in the developing zebrafish forebrain. J Vis Exp (50): 2594.
- Dong, Z., Yang, N., Yeo, S. Y., Chitnis, A. and Guo, S. (2012). Intralineage directional Notch signaling regulates self-renewal and differentiation of asymmetrically dividing radial glia. Neuron 74(1): 65-78.
- Fishell, G. and Kriegstein, A. R. (2003). Neurons from radial glia: the consequences of asymmetric inheritance. Curr Opin Neurobiol 13(1): 34-41.
- Gaiano, N. and Fishell, G. (2002). The role of notch in promoting glial and neural stem cell fates. Annu Rev Neurosci 25: 471-490.
- Horvitz, H. R. and Herskowitz, I. (1992). Mechanisms of asymmetric cell division: two Bs or not two Bs, that is the question. Cell 68(2): 237-255.
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B. and Schilling, T. F. (1995). Stages of embryonic development of the zebrafish. Dev Dyn 203(3): 253-310.
- Knoblich, J. A., (2001). Asymmetric cell division during animal development. Nat Rev Mol Cell Biol 2(1): 11-20.
- Kressmann, S., Campos, C., Castanon, I., Furthauer, M. and Gonzalez-Gaitan, M. (2015). Directional Notch trafficking in Sara endosomes during asymmetric cell division in the spinal cord. Nat Cell Biol 17(3): 333-339.
- Manders, E., Verbeek, F. J. and Aten, J. A. (1993). Measurement of co-localization of objects in dual-colour confocal images. J Microsc 169(3): 375-382.
- Mizutani, K., Yoon, K., Dang, L., Tokunaga, A. and Gaiano, N. (2007). Differential Notch signalling distinguishes neural stem cells from intermediate progenitors. Nature 449(7160): 351-355.
- Satoh, D., Sato, D., Tsuyama, T., Saito, M., Ohkura, H., Rolls, M. M., Ishikawa, F. and Uemura, T. (2008). Spatial control of branching within dendritic arbors by dynein-dependent transport of Rab5-endosomes. Nat Cell Biol 10: 1164-1171.
- Schindelin, J., Arganda-Carreras, I., Frise, E., Kaynig, V., Longair, M., Pietzsch, T., Preibisch, S., Rueden, C., Saalfeld, S. and Schmid, B. (2012). Fiji: an open-source platform for biological-image analysis. Nat Methods 9(7): 676-682.
- Shi, X., Li, Q., Dai, Z., Tran, A. A., Feng, S., Ramirez, A. D., Lin, Z., Wang, X., Chow, T. T. and Chen, J. (2021). Label-retention expansion microscopy. J Cell Biol 220(9): e202105067.
- Tong, K., Wagle, M. and Guo, S. (2019). Antibody Uptake Assay in the Embryonic Zebrafish Forebrain to Study Notch Signaling Dynamics in Neural Progenitor Cells In Vivo. Methods Mol Biol 1576: 273-281.
- Williams, S. E., Beronja, S., Pasolli, H. A. and Fuchs, E. (2011). Asymmetric cell divisions promote Notch-dependent epidermal differentiation. Nature 470(7334): 353-358.
- Zhao, X., Garcia, J. Q., Tong, K., Chen, X., Yang, B., Li, Q., Dai, Z., Shi, X., Seiple, I. B. and Huang, B. (2021). Polarized endosome dynamics engage cytoplasmic Par-3 that recruits dynein during asymmetric cell division. Sci Adv 7(24): eabg1244.
Article Information
Copyright
© 2022 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Readers should cite both the Bio-protocol article and the original research article where this protocol was used:
- Zhao, X., Garcia, J., Royer, L. A. and Guo, S. (2022). Colocalization Analysis for Cryosectioned and Immunostained Tissue Samples with or without Label Retention Expansion Microscopy (LR-ExM) by JACoP. Bio-protocol 12(5): e4336. DOI: 10.21769/BioProtoc.4336.
- Zhao, X., Garcia, J. Q., Tong, K., Chen, X., Yang, B., Li, Q., Dai, Z., Shi, X., Seiple, I. B. and Huang, B. (2021). Polarized endosome dynamics engage cytoplasmic Par-3 that recruits dynein during asymmetric cell division. Sci Adv 7(24): eabg1244.
Category
Cell Biology > Cell imaging > Fluorescence
Neuroscience > Development > Immunofluorescence
Molecular Biology > Protein > Protein-protein interaction
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.