- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Quantifying Lysosomal Degradation of Extracellular Proteins With a Fluorescent Protein-Based Internalization Assay
(*contributed equally to this work) Published: Vol 16, Iss 5, Mar 5, 2026 DOI: 10.21769/BioProtoc.5619 Views: 117
Reviewed by: David PaulAnonymous reviewer(s)
Original research article
The authors used this protocol in:

Mar 2023
Advertisement


Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Related protocols

Abstract
Endocytosis is an essential membrane transport mechanism that is indispensable for the maintenance of life. It is responsible for the selective internalization and subsequent degradation or recycling of specific extracellular proteins and nutrients, thereby facilitating cellular nutrient supply, modulation of receptor signaling, and clearance of foreign substances. However, methods for the quantitative analysis of lysosomal degradation of extracellular proteins via endocytosis remain limited. This protocol describes a method for purifying the protein-of-interest (POI)–red fluorescent protein (RFP)–green fluorescent protein (GFP) fusion protein, which is modified with specific mammalian cell glycans or other modifications, from the conditioned medium of mammalian cell cultures. Subsequently, the protocol details a quantitative approach for evaluating its internalization and lysosomal degradation within cells using the RFP–GFP tandem fluorescent reporter. Following the addition of POI-RFP-GFP to the medium, cells can be subjected to cell biological assays, such as flow cytometry, as well as biochemical analyses, such as immunoblotting. This protocol is broadly applicable to studies of the internalization of extracellular proteins.
Key features
• Purification of secreted GFP-RFP-fused POI from mammalian cell culture supernatant.
• Quantification of POI-RFP-GFP internalization through measurement of GFP and RFP signals using flow cytometry.
• Confirmation of lysosomal degradation of POI-RFP-GFP by immunoblotting.
Keywords: LysosomeGraphical overview
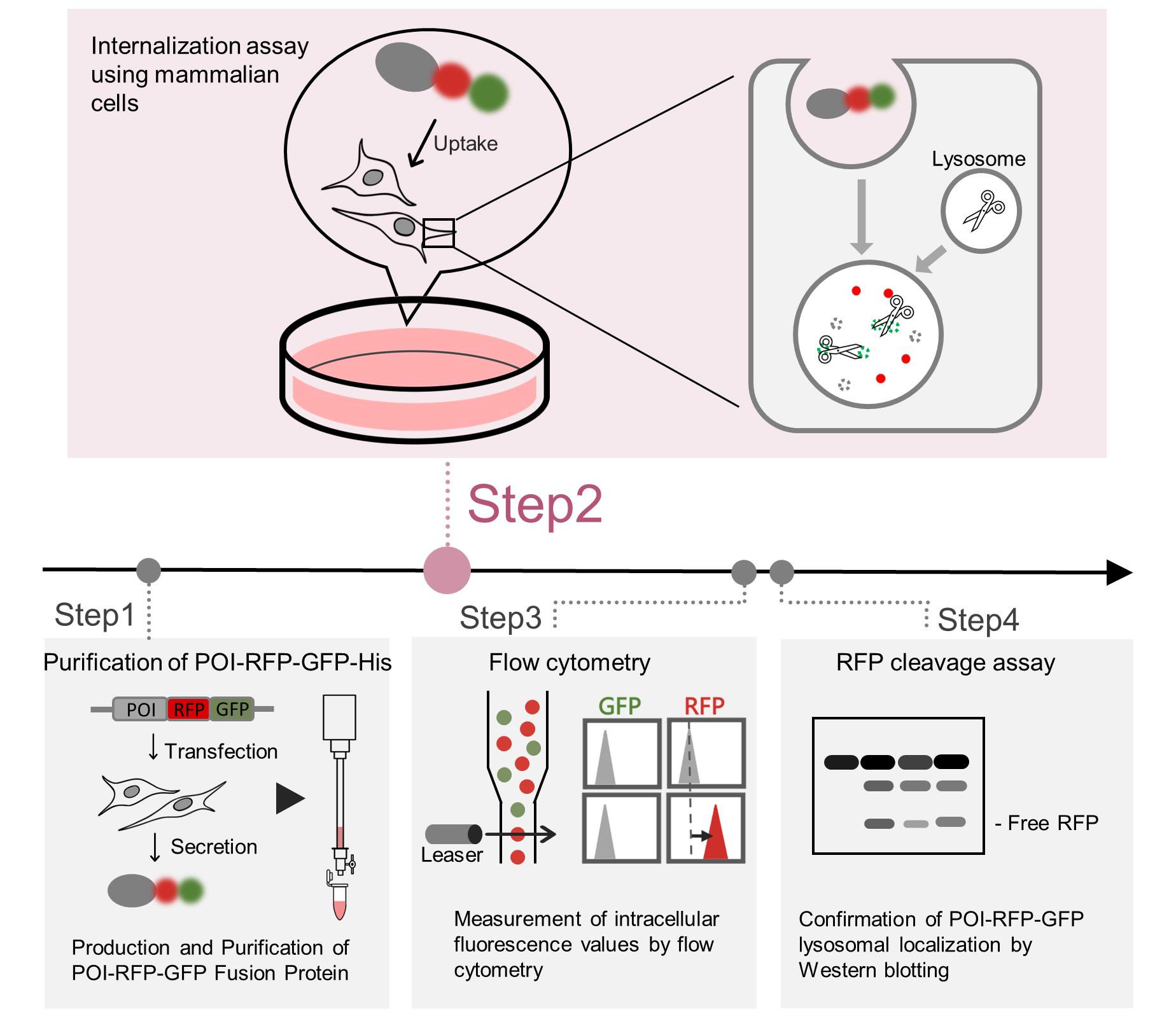
Background
Extracellular proteins, including those circulating in the bloodstream, are continuously secreted, internalized by mammalian cells, and subsequently degraded in lysosomes [1,2]. Proteins such as transferrin and immunoglobulins are recycled via the endosomes, whereas lipoproteins are constitutively degraded within lysosomes [3,4]. In contrast, various diseases, for example, Alzheimer’s disease, are associated with abnormal accumulation of extracellular proteins [5–7]. Accurate quantification of internalization is therefore essential in understanding extracellular proteostasis, pathological protein accumulation, and chaperone-dependent internalization pathways. However, methods for quantitatively tracking extracellular protein internalization and lysosomal degradation have long been limited, posing significant challenges for such analyses. Covalent fluorescent labeling approaches rely on random conjugation of fluorophores to proteins of interest (POIs), which can interfere with protein structure or function. Moreover, increases in cellular fluorescence often fail to distinguish whether the signal reflects simple binding of the POI to the cell surface or enhanced internalization. The differences between the red fluorescent protein (RFP)–green fluorescent protein (GFP) fusion system and covalent fluorescent labeling are summarized in Table 1.
Tandem RFP-GFP reporters have traditionally been used to monitor intracellular lysosomal transport processes, such as autophagy [8]. We have, for the first time, adapted this system to monitor the lysosomal degradation of extracellular proteins. The internalization reporter system, in which a POI is fused to an RFP–GFP tandem fluorescent tag, was specifically developed to quantify this extracellular protein endocytosis and lysosomal transport [9–11]. GFP is quenched in the acidic environment of lysosomes, whereas RFP retains fluorescence; thus, GFP serves as an indicator of cell-surface binding or internalized but not yet transported to lysosomes, while an increase in RFP reflects accumulation at both the cell surface and within lysosomes. Therefore, an increase in RFP fluorescence without a corresponding increase in GFP fluorescence indicates that the POI-RFP-GFP fusion protein has undergone lysosomal degradation. This reporter enables sensitive, quantitative, and spatial analysis of both internalization and lysosomal transport of the POI.
This protocol describes an internalization assay using POI-RFP-GFP fusion proteins and presents a comprehensive workflow that integrates protein purification, flow cytometry, and immunoblotting. Together, these approaches enable robust evaluation of lysosomal degradation of extracellular protein.
The protocol has been used in Tomihari et al. (2023) [9], where it contributed to elucidating the mechanisms of extracellular protein internalization and lysosomal degradation.
Table 1. Comparison of conventional methods and the red fluorescent protein (RFP)–green fluorescent protein (GFP) tandem reporter
| Method | Principle | Advantages | Limitations | Target detection |
|---|---|---|---|---|
Covalent fluorescence labeling | Chemical conjugation of fluorescent dyes to Lys/Cys residues | Applicable to numerous proteins | Random labeling may interfere with protein function; increased fluorescence does not always reflect internalization | Cell surface and lysosome (indistinguishable by flow cytometry; distinguishable by microscopy) |
| GFP-only reporter | Fusion of GFP tag to the N- or C-terminus of POI | Smaller molecular weight than the tandem reporter | Since GFP is degraded in the lysosomes, lysosomal transport cannot be detected via the GFP signal | Cell surface |
| RFP-only reporter | Fusion of RFP tag to the N- or C-terminus of POI | Smaller molecular weight than the tandem reporter; the RFP cleavage assay enables biochemical analysis of lysosomal transport | Cell surface binding and lysosomal transport are indistinguishable by flow cytometry | Cell surface and lysosome (indistinguishable by flow cytometry; distinguishable by RFP cleavage assay and microscopy) |
| RFP-GFP tandem reporter (current protocol) | Fusion of RFP-GFP tag to the N- or C-terminus of POI | Simultaneous evaluation of cell surface and lysosomal transport; the RFP cleavage assay enables biochemical analysis of lysosomal transport | Large tag may affect the function of certain POIs | Cell surface and lysosome (distinguishable by flow cytometry, RFP cleavage assay, and microscopy) |
Materials and reagents
Biological materials
1. Human: Flp-In T-REx 293 cell line (Thermo Fisher, catalog number: R78007)
2. Human: 293FT cell line (Thermo Fisher, catalog number: R70007)
3. Human: HeLa cells (ATCC, catalog number: CCL-2)
Plasmids
1. pcDNA5/FRT/TO vector (Thermo Fisher, catalog number: V601020)
2. pOG44 Flp-recombinase expression vector (Thermo Fisher, catalog number: V600520)
3. pcDNA5/FRT/TO–α2M–RG–His (RIKEN BioResource Research Center, catalog number: RDB20287)
Reagents
1. Dulbecco’s modified Eagle medium (DMEM), high glucose (Nacalai Tesque, catalog number: 16971-55)
2. Advanced DMEM/F-12 (Thermo Fisher, catalog number: 12634010)
Note: Reduced serum medium (DMEM/Ham’s F-12) (Nacalai Tesque, catalog number: 21906-55) is also compatible with this protocol.
3. Fetal bovine serum (FBS) (MERCK, catalog number: 12003C)
4. Penicillin-streptomycin (Nacalai Tesque, catalog number: 09367-34)
5. Opti-MEM (Thermo Fisher, catalog number: 31985070)
6. MiniPrep kit (Favorgen, catalog number: FAPDE 001-1)
7. Polyethylenimine (Polysciences, catalog number: 24765-2)
8. Hygromycin (Fujifilm, catalog number: 085-06153)
9. Doxycycline (Sigma, catalog number: D9891)
10. 2.5 g/L trypsin, 1 mmol/L EDTA solution (Nacalai Tesque, catalog number: 32777-44)
11. Bafilomycin A1 (Cayman, catalog number: 11038)
12. Ni-NTA agarose resin (Fujifilm, catalog number: 141-09683)
13. Imidazole (Fujifilm, catalog number: 099-00013)
14. Sodium chloride (NaCl) (Fujifilm, catalog number: 191-01665)
15. di-sodium hydrogenphosphate dodecahydrate (Na2HPO4·12H2O) (Nacalai Tesque, catalog number: 31722-45)
16. Potassium dihydrogenphosphate (KH2PO4) (Nacalai Tesque, catalog number: 28721-55)
17. Potassium chloride (KCl) (Kanto Kagaku, catalog number: 32326-00)
18. D-PBS without Ca and Mg, powder (Nacalai Tesque, catalog number: 07269-84)
19. Triton X-100 (Nacalai Tesque, catalog number: 35501-02)
20. Polyoxyethylene (20) sorbitan monolaurate (100% Tween20) (Fujifilm, catalog number: 166-21115)
21. Glycerol (Nacalai Tesque, catalog number: 17017-35)
22. 2-Mercaptoethanol (Fujifilm, catalog number: 198-15781)
23. Coomassie Brilliant Blue R-250 (Fujifilm, catalog number: 031-17922)
24. Skim milk (Nacalai Tesque, catalog number: 31149-75)
25. DAPI (Nacalai Tesque, catalog number: 11034-14)
26. Tris(hydroxymethyl)aminomethane (Tris) (Nacalai Tesque, catalog number: 35406-91)
27. Methanol (Nacalai Tesque, catalog number: 21915-93)
28. Acetic acid (Nacalai Tesque, catalog number: 00212-43)
29. Hydrochloric acid (HCl) (Fujifilm, catalog number: 080-01066)
30. 100× protease inhibitor cocktail (Nacalai Tesque, catalog number: 03969-21)
31. Phenylmethanesulfonyl fluoride (PMSF) (MP Biomedicals, catalog number: MPB195381-5)
32. Signal enhancer HIKARI for Western Blotting and ELISA (Nacalai Tesque, catalog number: 02267-41)
33. Rabbit polyclonal anti-mCherry (homemade), mouse monoclonal anti-RFP (MBL, catalog number: M204-3), or other mCherry antibodies
34. Mouse monoclonal anti-α tubulin (Fujifilm , catalog number: 071-25031)
35. Anti-mouse IgG, HRP-linked antibody (CST, catalog number: 7076S)
36. Anti-rabbit IgG, HRP-linked antibody (CST, catalog number: 7074S)
37. Recombinant luciferase (Promega, catalog number: E1702)
38. Purified recombinant His-Alfa-S-formylglutathione hydrolase (ESD) (homemade; see [9])
39. Purified recombinant Clusterin-mCherry-sfGFP-His (homemade; see [10])
Solutions
1. Column wash buffer (see Recipes)
2. Column elution buffer (see Recipes)
3. DMEM complete medium (see Recipes)
4. Trypsin solution (see Recipes)
5. FCM buffer (see Recipes)
6. 10× lysis buffer (see Recipes)
7. Lysis buffer (see Recipes)
8. 1 M imidazole (see Recipes)
9. Coomassie Brilliant Blue (CBB) stain solution (see Recipes)
10. Coomassie Brilliant Blue (CBB) bleaching (see Recipes)
11. 25× TBS (see Recipes)
12. 5% Tween20 (see Recipes)
13. TBST (see Recipes)
14. 25× PBS (see Recipes)
15. PBS (see Recipes)
Recipes
1. Column wash buffer
| Reagent | Final concentration | Quantity |
|---|---|---|
| 5 M NaCl | 500 mM | 100 mL |
| 1 M Imidazole | 10 mM | 10 mL |
| 25× PBS | 1× | 40 mL |
| H2O | N/A | 850 mL |
| Total | N/A | 1,000 mL |
Store at 4 °C.
2. Column elution buffer
| Reagent | Final concentration | Quantity |
|---|---|---|
| 5 M NaCl | 150 mM | 12 mL |
| 1 M Imidazole | 200 mM | 80 mL |
| 25× PBS | 1× | 16 mL |
| H2O | N/A | 292 mL |
| Total | N/A | 400 mL |
Store at 4 °C.
3. DMEM complete medium
| Reagent | Final concentration | Quantity |
|---|---|---|
| DMEM | N/A | 500 mL |
| FBS | 10% | 55 mL |
| Penicillin-streptomycin | 1% | 5.5 mL |
| Total | N/A | 555 mL |
Store at 4 °C.
4. Trypsin solution
| Reagent | Final concentration | Quantity |
|---|---|---|
| 2.5 g/L trypsin and 1 mmol/L EDTA solution | 0.5 g/L trypsin and 0.2 mmol/L EDTA | 10 mL |
| PBS | N/A | 40 mL |
| Total | N/A | 50 mL |
Store at 4 °C.
5. FCM buffer
| Reagent | Final concentration | Quantity |
|---|---|---|
| FBS | 5% | 750 μL |
| 1 mg/mL DAPI | 1 μg/mL | 15 μL |
| PBS | N/A | 14.25 mL |
| Total | N/A | 15 mL |
Store at 4 °C. Prepare immediately before use.
6. 10× lysis buffer
| Reagent | Final concentration | Quantity |
|---|---|---|
| 1 M Tris-HCl pH 7.5 | 500 mM | 50 mL |
| 5 M NaCl | 1500 mM | 30 mL |
| 0.5 M EDTA pH 8.0 | 10 mM | 2 mL |
| Triton X-100 | 10% | 10 mL |
| H2O | N/A | 8 mL |
| Total | N/A | 100 mL |
Store at 4 °C.
7. Lysis buffer
| Reagent | Final concentration | Quantity |
|---|---|---|
| 10× lysis buffer | 1× | 100 μL |
| 100× protease inhibitor cocktail | 1× | 10 μL |
| 0.1 M PMSF | 1 mM | 10 μL |
| H2O | N/A | 880 μL |
| Total | N/A | 1,000 μL |
Store at 4 °C. Prepare immediately before use.
8. 1 M imidazole
1 M imidazole in H2O, with pH adjusted to 8.0 using HCl. Store at 20 °C.
9. Coomassie Brilliant Blue (CBB) stain solution
| Reagent | Final concentration | Quantity |
| Coomassie Brilliant Blue R-250 | 0.1% | 1 g |
| Methanol | 40% | 400 mL |
| Acetic acid | 10% | 100 mL |
| H2O | N/A | 500 mL |
| Total | N/A | 1,000 mL |
Store at 20 °C.
10. Coomassie Brilliant Blue (CBB) bleaching solution
| Reagent | Final concentration | Quantity |
| Methanol | 40% | 400 mL |
| Acetic acid | 10% | 100 mL |
| H2O | N/A | 500 mL |
| Total | N/A | 1,000 mL |
Store at 20 °C.
11. 25× TBS
| Reagent | Final concentration | Quantity |
| Tris | 0.5 M | 605 g |
| NaCl | 3.42 M | 2,000 g |
| H2O | N/A | Bring the final volume to 10 L |
Adjust the pH to 7.6 with HCl. Store at 20 °C.
12. 5% Tween 20
| Reagent | Final concentration | Quantity |
| 100% Tween20 | 5% | 25 mL |
| H2O | N/A | 475 mL |
| Total | N/A | 500 mL |
Pipette until well mixed, then autoclave. Store at 20 °C.
13. TBST
| Reagent | Final concentration | Quantity |
| 25× TBS | 1× | 100 mL |
| 5% Tween20 | 0.02% | 50 mL |
| H2O | N/A | 2,350 mL |
| Total | N/A | 2,500 mL |
Store at 20 °C.
14. 25× PBS
| Reagent | Final concentration | Quantity |
| NaCl | 3.42 M | 1,000 g |
| Na2HPO4·12H2O | 0.2 M | 362.5 g |
| KH2PO4 | 0.037 M | 25 g |
| KCl | 0.067 M | 25 g |
| H2O | N/A | Bring the final volume to 5 L |
Store at 20 °C.
15. PBS
| Reagent | Final concentration | Quantity |
| D-PBS without Ca and Mg, Powder | N/A | 48 g |
| H2O | N/A | Bring the final volume to 5 L |
After mixing with a stirrer until dissolved, autoclave. Store at 20 °C.
Laboratory supplies
1. 60-mm dish (VIOLAMO, catalog number: 2-8590-02)
2. 100-mm dish (VIOLAMO, catalog number: 2-8590-03)
3. 12-well plates (VIOLAMO, catalog number: 2-8588-02)
4. 24-well plates (VIOLAMO, catalog number: 2-8588-03)
5. Econo-column chromatography columns, 0.7 × 10 cm (Bio-Rad, catalog number: 7370712B02)
6. Econo-column funnels (Bio-Rad, catalog number: 7310003)
7. Microsep Advance Centrifugal Devices with Omega Membrane 30 K (PALL, catalog number: MCP030C46)
8. Easy strainer 70 μm (Greiner, catalog number: 542070)
Equipment
1. Flow cytometer (Beckman Coulter, model: CytoFLEX S)
Procedure
A. Generation of Flp-In T-REx 293 cells stably expressing POI-RFP-GFP-(6×)His
Notes:
1. A stable cell line is generated using the Flp-In system, in which the Flp recombinase target (FRT) site in the pcDNA5/FRT/TO vector is integrated into the genome of Flp-In 293 cells [12]. This system enables robust and uniform high-level expression, even for large fusion proteins. Alternative expression methods, including viral vectors and transient transfection, can also be used, provided that the POI is highly overexpressed.
2. The T-REx (Tet-on) inducible system is not essential for expressing POI-GFP-RFP; however, it can be advantageous when strong overexpression of the POI causes cytotoxicity.
3. mCherry, an acid- and protease-resistant fluorescent protein, is used as the RFP [13]. sfGFP is pH-sensitive, monomeric, resistant to disulfide bond formation, and lacks N-glycosylation sites [14]; therefore, mCherry and sfGFP are recommended for fusion with secreted proteins. While other fluorescent proteins can be utilized, caution is advised when using cysteine-containing fluorescent proteins, as they may form undesirable disulfide bonds in the endoplasmic reticulum, thereby reducing secretion efficiency.
4. If the structural information or epitope tag compatibility of the POI is unknown, it is recommended to test both POI-RFP-GFP and RFP-GFP-POI constructs.
Critical: If the POI is a secreted protein, replace its native signal sequence with the signal sequence located at the N-terminus of 6× His-RFP-GFP-POI.
1. Vector construction: Subclone POI-RFP-GFP-(6×)His or SS (signal sequence)–(6×)His-RFP-GFP-POI into the pcDNA5/FRT/TO vector. In this protocol, α2-macroglobulin (α2M)-RFP-GFP-(6×)His (α2M-RG-His) is used.
a. Primers were designed specifically for Gibson Assembly to amplify the sequences of RFP, GFP, 6×His, the protein of interest (POI), and the vector backbone [15].
b. The resulting PCR products were purified and subsequently incubated with a reaction mixture containing T5 exonuclease, Phusion DNA polymerase, and Taq DNA ligase.
c. The assembled products were transformed into competent E. coli cells. Positive clones were screened, and the plasmids were isolated using a MiniPrep kit to obtain the target DNA constructs.
2. Introduction into Flp-In T-REx 293 cells (co-transfection)
a. Seed Flp-In T-REx 293 cells in a 12-well plate and culture until 30%–50% confluence.
b. Prepare 2 μL polyethylenimine MAX in 100 μL of Opti-MEM (solution A) and incubate at room temperature for at least 5 min.
c. Prepare 0.4 μg of pcDNA5/FRT/TO–α2M–RG–His plus 0.4 μg of pOG44 in 100 μL of Opti-MEM (solution B).
d. Add solution B to solution A, mix gently, and incubate for 20 min at room temperature.
Note: Incubation up to approximately 60 min is acceptable.
e. Aspirate the culture medium, replace the medium of T-REx cells with 0.75 mL per well of DMEM containing 10% FBS (without penicillin and streptomycin), and then add 200 μL of the transfection mixture. Culture for 4–12 h at 37 °C with 5% CO2.
Note: Overnight incubation is not recommended, as it may cause cell death.
f. Aspirate the culture medium, replace with 1 mL of regular medium (DMEM containing 10% FBS and 1% penicillin-streptomycin), and culture for an additional 4–6 days.
3. Selection of stable clones
a. Replace the medium with DMEM containing 100 μg/mL hygromycin (depending on the vector used) and culture for 2–3 weeks.
Notes:
1. During the 2–3-week selection period, monitor the cells regularly. If the culture reaches confluence, split the cells at a ratio of 1:4 to 1:8. Additionally, if significant cell debris or dead cells accumulate, replace the medium with fresh selection medium to optimize the growth environment for the hygromycin-resistant clones.
2. Since the Flp-In 293 system ensures that all selected cells are high-expression clones due to site-specific integration, single-cell cloning is not required. However, when generating stable cell lines using other transfection methods, it is advisable to perform single-cell cloning to select for high-expression clones.
B. Purification of secreted POI (α2M)-RFP-GFP-His from the conditioned medium
1. Scale up the stable cell line generated in section A by seeding into four 100 mm dishes and subsequently expand the culture to twenty 100 mm dishes using regular medium supplemented with 0.1 μg/mL doxycycline. Culture for several days until cells reach approximately 80% confluence.
2. Aspirate the culture medium and wash each dish once with 3 mL of PBS.
3. Remove PBS and replace with 10 mL per dish of advanced DMEM/F-12 (without serum) containing 0.1 μg/mL doxycycline. Culture for 4 days.
Critical: Do not add FBS to the medium, as it can cause nonspecific protein contamination during purification.
Notes:
1. If cells remain healthy on day 4, extending the culture to 5–6 days can increase protein yield. However, excessive accumulation of dead cells significantly elevates the risk of cytosolic protein contamination.
2. To confirm that the selected cell population is producing the tandem-tagged POI, observe the culture medium under a fluorescence microscope to check for red or green fluorescence.
4. Collect the conditioned medium into four 50-mL tubes.
5. Centrifuge at 2,300–4,000× g for 10 min at 4 °C to remove cell debris.
6. Transfer the supernatant to a sterile 200-mL bottle and store at -80 °C.
Critical: Protect samples from light to prevent photobleaching.
Pause point: Conditioned medium can be stored at -80 °C for several years.
7. Thaw the -80 °C conditioned medium.
8. Pack 1 mL (bed volume) of Ni-NTA agarose resin into a gravity-flow column (Figure 1).
9. Equilibrate the resin with 10 mL of column wash buffer.
10. Add imidazole to the thawed medium to a final concentration of 10 mM (e.g., add 2 mL of 1 M imidazole stock to 200 mL of medium).
Critical: Imidazole reduces nonspecific binding and improves purity.
Note: For SDS-PAGE analysis, collect 20 μL into a 1.5-mL tube and store at 4 °C.
11. Load the medium onto the gravity-flow column. Slowly load the initial few milliliters until the column dead volume is cleared, then load the remaining medium (Figure 1).
Notes:
1. Flow rate is approximately 1–2 mL/min.
2. Collect 20 μL of flowthrough for SDS-PAGE analysis and store at 4 °C.
12. Wash the column with wash buffer to remove nonspecifically bound proteins: add 1 mL first, followed by four subsequent additions of 10 mL each.
13. Elute the target protein using column elution buffer and collect three fractions: 600 μL (Fraction 1), 2,000 μL (Fraction 2), and 1,000 μL (Fraction 3) (Figure 1).
Note: The target protein primarily elutes in Fraction 2, which appears red due to the inherent color of the RFP-GFP.
14. Measure absorbance at 280 nm (A280) for each fraction and identify the fraction with the highest protein content (typically Fraction 2).
Note: For SDS-PAGE, collect 3 μL of each fraction (Fractions 1–3) and store at 4 °C.
15. Concentrate the target protein and remove imidazole by ultrafiltration:
a. Transfer the selected fraction to a Microsep Advance centrifugal device (30 kDa MWCO; the cutoff should be one-third or less of the target protein’s molecular weight) and centrifuge at 2,380× g for 30 min at 4 °C until the volume is reduced to 0.2 mL.
b. Discard the flowthrough and add 2 mL of PBS to the device.
c. Repeat steps B15a–b at least three times.
d. Transfer the concentrated protein solution to a new 1.5-mL tube.
Critical: More than 2 mM imidazole in the culture medium inhibits lysosomal activity [16]; therefore, imidazole must be removed from purified proteins prior to the internalization assay.
Optional: Imidazole can also be removed by dialysis.
16. Add glycerol to a final concentration of 10%–20% and mix thoroughly.
Critical: Glycerol prevents aggregation during freeze–thaw cycles. It can be omitted if it impairs protein functionality.
17. Measure protein concentration at A280 and store at -80 °C.
Notes:
1. Typically, 1–4 mg of protein is obtained from 200 mL of conditioned medium.
2. Aliquot into small volumes (10–50 μL) to minimize freeze–thaw cycles.
18. Mix a portion of the purified protein, conditioned medium, or flowthrough with 2× SDS sample buffer containing 10% 2-mercaptoethanol and boil at 95 °C for 5 min.
19. Run the samples on SDS-PAGE and stain with CBB solution to confirm protein purity and molecular weight (Figure 2).
Note: CBB is a widely used dye that visualizes protein bands after SDS-PAGE by binding to proteins via electrostatic and hydrophobic interactions [17].

Figure 1. Purification of α2M-RFP-GFP-His using Ni-NTA agarose resin. (A) The Ni-NTA column is assembled and packed with equilibrated resin. (B) The conditioned medium containing α2M-RFP-GFP-His is applied to the column. (C) After binding, the resin becomes visibly red due to the retained α2M-RFP-GFP-His. (D) During elution with elution buffer, the resin gradually loses its red color, indicating the release of the bound protein. (E) The eluate fractions collected in microtubes contain the purified POI-RFP-GFP-His.
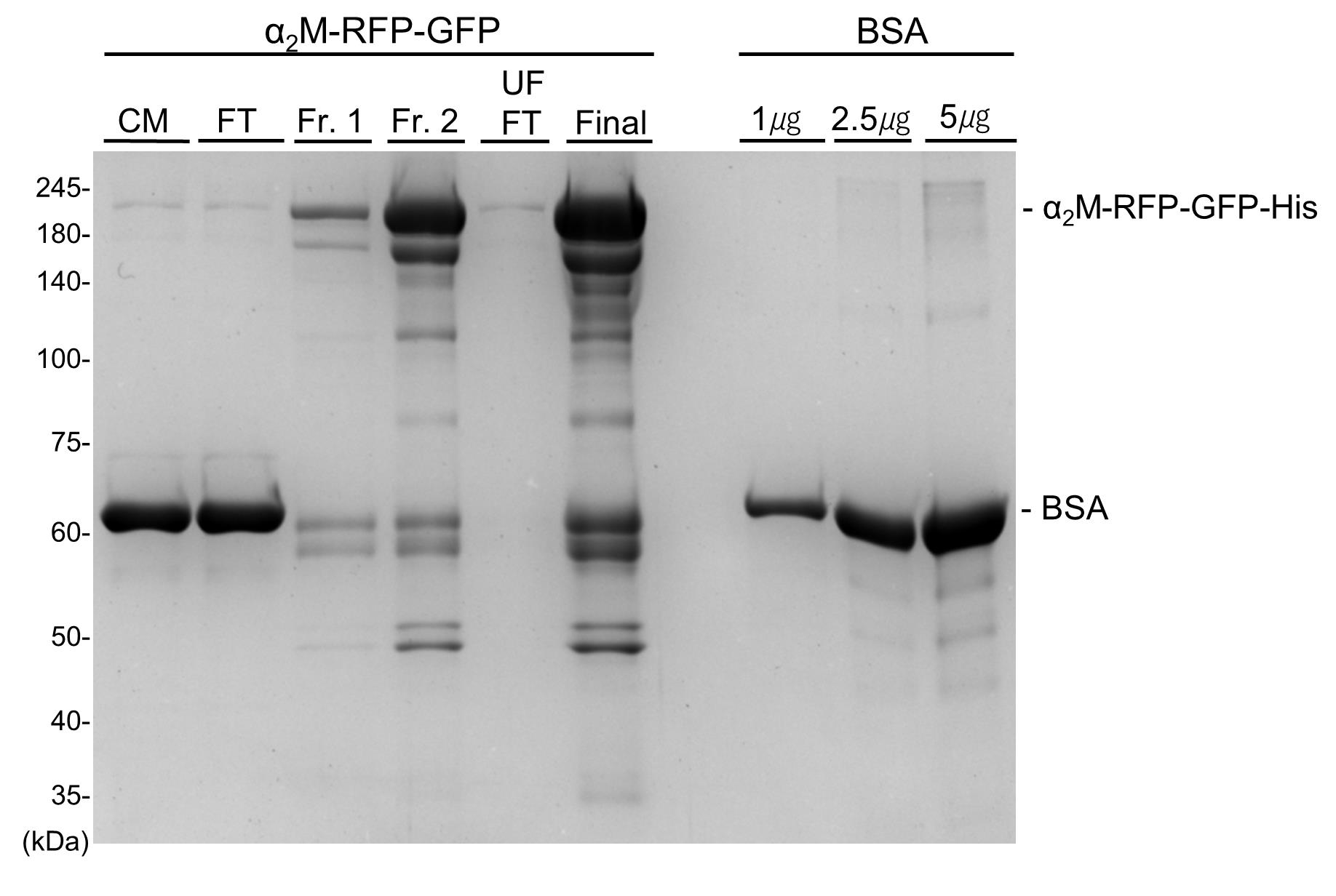
Figure 2. Confirmation of Coomassie Brilliant Blue (CBB) staining for purified α2M-RFP-GFP-His protein. Purification of α2M-RFP-GFP-His from conditioned medium of mammalian cells. CBB staining of α2M-RFP-GFP-His. Samples are subjected to SDS-PAGE followed by CBB staining. CM: conditioned medium (input sample); FT: flowthrough; Fr. 1: elution fraction 1; Fr. 2: elution fraction 2; UF FT: ultrafiltration flowthrough; Final: purified protein.
C. Flow cytometric analysis of internalization using RFP-GFP tandem reporter proteins
Day 1: Cell seeding
1. Seed HeLa cells at 30%–50% confluence (or a relevant cell line for the POI) into a 24-well plate.
Day 2: Sample preparation and treatment
2. Prepare the reagents to be added to cells.
a. Add purified α2M-RFP-GFP-His (final 0.2 μM) to advanced DMEM/F-12 (250 μL).
Optional: To generate misfolded α2M substrate, add His-Alfa-S-formylglutathione hydrolase (ESD) to a final concentration of 8 μM and apply heat stress. Incubate the medium containing ESD and α2M-RFP-GFP-His at 50 °C for 1 h.
Notes:
1. ESD is an endoplasmic reticulum (ER) protein that readily misfolds and binds to α2M-RG upon exposure to heat stress at 50 °C for 1 h, and the resulting α2M-RG-ESD complex is subsequently transported to the lysosomes [9].
2. The optimal concentration of the POI is protein-dependent. It may be useful to test a concentration range between 0.1 and 1 μM.
3. Ensure that equal amounts of POI-RG are added to each treatment medium. Inconsistency in the initial POI-RG concentration across samples can lead to substantial variability in the final readout.
3. Wash the cells once with serum-free DMEM, replace with the medium containing α2M-RFP-GFP-His, and incubate for 16 h at 37 °C with 5% CO2.
Note: Avoid overconfluence as it reduces internalization efficiency.
Day 3: Cell collection and flow cytometry
4. Cell collection
a. Wash each well of the 24-well plate with PBS.
b. Remove PBS and add 100 μL/well of trypsin. Incubate for 5 min at 37 °C.
c. Add 200 μL of FCM buffer and pipette to resuspend the cells.
d. Filter the cell suspension through a cell strainer and collect it into a 1.5-mL FCM tube.
e. Store the samples on ice, protected from light, until analysis.
5. Flow cytometry
a. Turn on the flow cytometer and run the start-up program.
Note: Follow the manufacturer’s instructions for the start-up procedure.
b. Load the sample tubes and begin acquisition.
i. Set FSC-A vs. SSC-A to gate the main cell population and exclude debris.
ii. Use SSC-H vs. SSC-W to gate single cells and exclude doublets.
iii. Use FSC-A vs. DAPI-A to gate viable cells and exclude dead cells.
iv. Acquire 10,000 cells from the final gated population.
v. Detect sfGFP using a 525/40-nm bandpass filter with 488-nm excitation and detect mCherry using a 585/42-nm bandpass filter with 561-nm excitation.
Note: Filter settings may vary among instruments; confirm the specifications for the system used.
D. Immunoblot analysis of RFP cleavage assay using RFP-GFP tandem reporters
Day 1: Cell seeding
1. Seed HeLa cells at 30%–50% confluence (or any POI-relevant cell line) into a 12-well plate.
Day 2: Sample preparation and treatment
2. Prepare the reagents to be added to the cells. Treat the cells with the POI-RFP-GFP fusion protein as described in steps C2 and C3.
3. Aspirate the medium containing POI-RFP-GFP after 16 h.
Day 3: Cell collection and lysis
4. Cell lysis
a. Wash each well of the 12-well plate with PBS.
b. Remove PBS and add 150 μL/well of trypsin. Incubate for 5 min at 37 °C.
c. Add 1 mL of cold PBS containing 10% FBS, resuspend thoroughly, and collect into a 1.5-mL tube.
Note: FBS is required to inhibit trypsin.
d. Centrifuge at 2,300× g for 2 min at 4 °C.
e. Discard the supernatant, resuspend the pellet in 1 mL of cold PBS, and centrifuge at 2,300× g for 2 min at 4 °C.
f. Discard the supernatant and lyse cells in 50 μL of lysis buffer.
g. Incubate for 15 min on ice.
h. Centrifuge at 20,600× g for 5 min at 4 °C to remove insoluble material.
i. Transfer the supernatant to a new 1.5-mL tube, dividing it into 40 μL and 10 μL portions.
i. Mix the 40-μL portion with 10 μL of 6× sample buffer and boil at 95 °C for 5 min.
ii. Use the 10-μL portion for protein quantification by Bradford assay.
5. SDS-PAGE and immunoblotting
a. Load 10–20 μg of boiled samples per lane and run SDS-PAGE for 90 min.
b. Transfer proteins to a PVDF membrane for 1 h.
c. Block with 5% skim milk for 30 min at room temperature.
d. Incubate with primary antibodies—rabbit anti-RFP (1:2,000) and mouse anti-β-tubulin (1:2,000)—for 1 h at room temperature.
Note: The primary antibody was diluted with HIKARI A solution (Nacalai Tesque).
e. Wash the membrane three times for 5 min each with TBST.
f. Incubate with HRP-conjugated secondary antibodies—anti-mouse IgG (1:2,000)—for 1 h at room temperature.
Note: The secondary antibody was diluted with HIKARI B solution (Nacalai Tesque).
g. Wash the membrane three times for 5 min each with TBST.
h. Incubate the membrane with ImmunoStarR Zeta HRP substrate for 5 min.
i. Place the membrane in the Fusion SOLO chamber and acquire images.
Data analysis
A. Evaluation of intracellular internalization of α2M-RFP-GFP-His by flow cytometry
In lysosomes, GFP is inactivated, whereas RFP is highly resistant to both acidic pH and lysosomal proteases. By leveraging this difference in stability, a reporter in which an RFP-GFP tandem tag is fused to a POI enables evaluation of whether the POI has been transported to lysosomes, based on changes in fluorescence intensities. During intracellular transport, the GFP signal decreases due to the lysosomal environment, whereas the RFP signal remains stable. Therefore, an increase in the RFP signal alone indicates that the POI has been delivered to lysosomes. In contrast, simultaneous increases in both the GFP and RFP imply that the POI is retained at the cell surface or in early endosomal compartments. Because flow cytometry quantitatively measures GFP and RFP intensities at the single-cell level, it allows robust and highly quantitative analysis of POI internalization and degradation stage. The purpose of flow cytometry is to quantify red and green fluorescence. This allows evaluation of the efficiency of POI internalization and lysosomal transport. To confirm statistical significance, it is recommended that flow cytometry be performed in at least three independent experiments.
In the internalization assay for α2M-RFP-GFP-His described in this protocol, the addition of heat-stressed ESD resulted in a marked increase in the RFP signal compared with when α2M-RFP-GFP-His was added alone (Figure 3). This implies that the presence of misfolded proteins promotes the internalization and lysosomal degradation of α2M. Although RFP intensity was also observed when α2M-RFP-GFP-His alone was added, this signal was attributed to nonspecific uptake of extracellular proteins via fluid-phase endocytosis. Furthermore, because bafilomycin A1 inhibits both lysosomal acidification and endocytosis, bafilomycin A1 treatment resulted in suppression of RFP intensity and a simultaneous increase in GFP intensity.
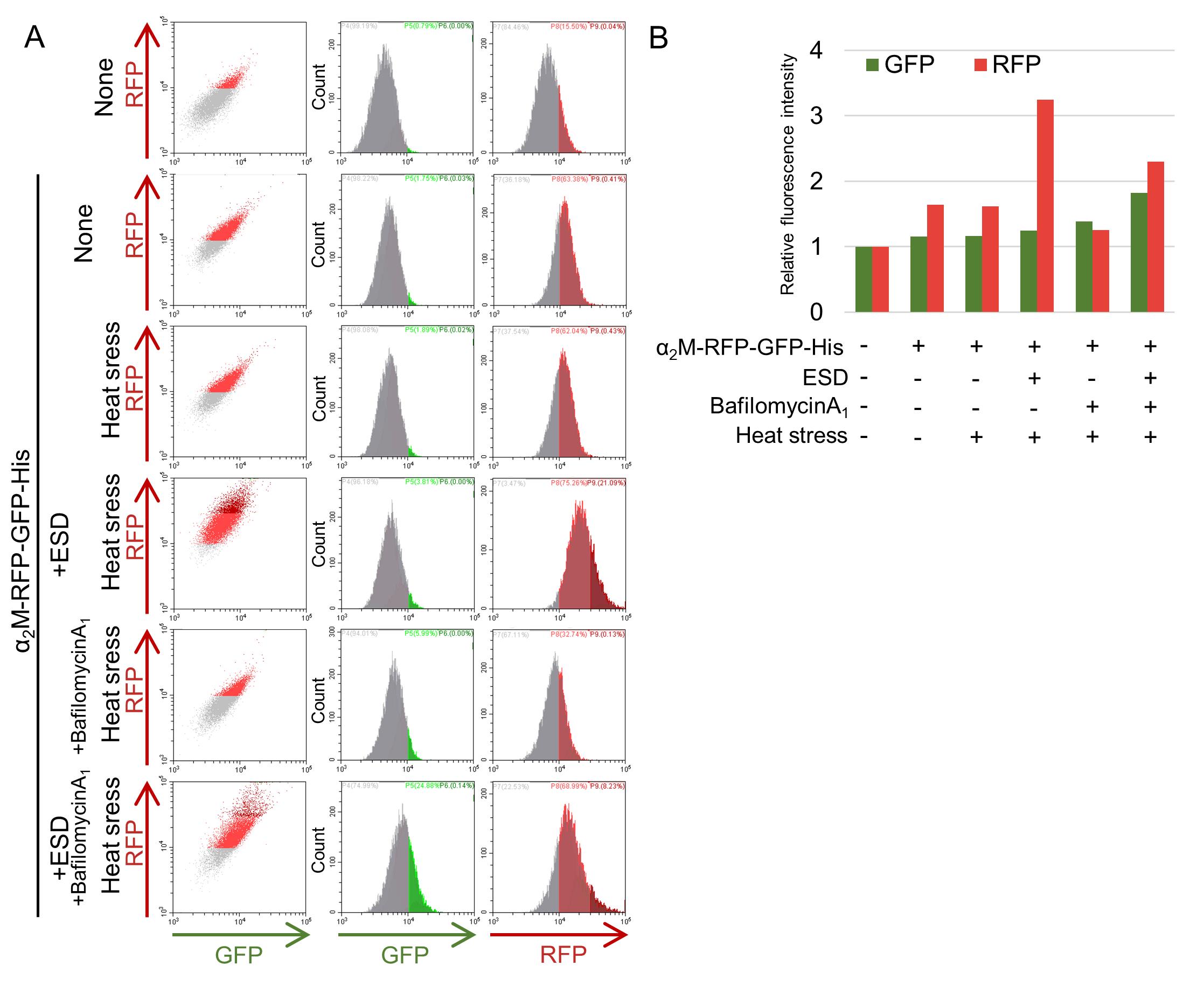
Figure 3. Analysis of α2M-RFP-GFP-His internalization by flow cytometry. Purified α2M-RFP-GFP-His was added to reduced-serum DMEM/F-12 either alone or together with ESD and preincubated at room temperature or at 50 °C (heat stress) for 1 h. HeLa cells were then cultured in medium with or without bafilomycin A1 for 16 h at 37 °C, followed by analysis using flow cytometry. (A) Results from flow cytometric analysis. (B) Graph illustrating the relative fluorescence intensities of GFP and RFP.
B. Analysis of RFP-cleavage assay by immunoblotting
By exploiting the high stability of RFP against lysosomal proteases, intracellular internalization of POI-RFP-GFP-His can also be assessed biochemically via immunoblotting. When α2M-RFP-GFP-His is transported to lysosomes, α2M and GFP are degraded, whereas RFP remains stable even after cleavage. Therefore, detection of cleaved RFP accumulating in lysosomes serves as a direct indicator that the POI has undergone lysosomal degradation.
Addition of α2M-RFP-GFP-His alone resulted in increased intracellular levels of cleaved RFP (Figure 4). When heat-stressed ESD was co-administered, the amount of cleaved RFP increased further compared with α2M alone, implying that α2M complexed with misfolded proteins is selectively internalized and degraded in lysosomes. In contrast, treatment with bafilomycin A1 nearly completely suppressed the generation of cleaved RFP, leading to the accumulation of full-length α2M-RFP-GFP-His. These findings support the conclusion that cleaved RFP is a reliable marker of lysosome-dependent degradation.
Collectively, these results demonstrate that α2M-RFP-GFP-His was internalized by cells, trafficked to lysosomes, and subsequently degraded. The accumulation of cleaved RFP in immunoblotting provides a robust method for validating lysosomal degradation.
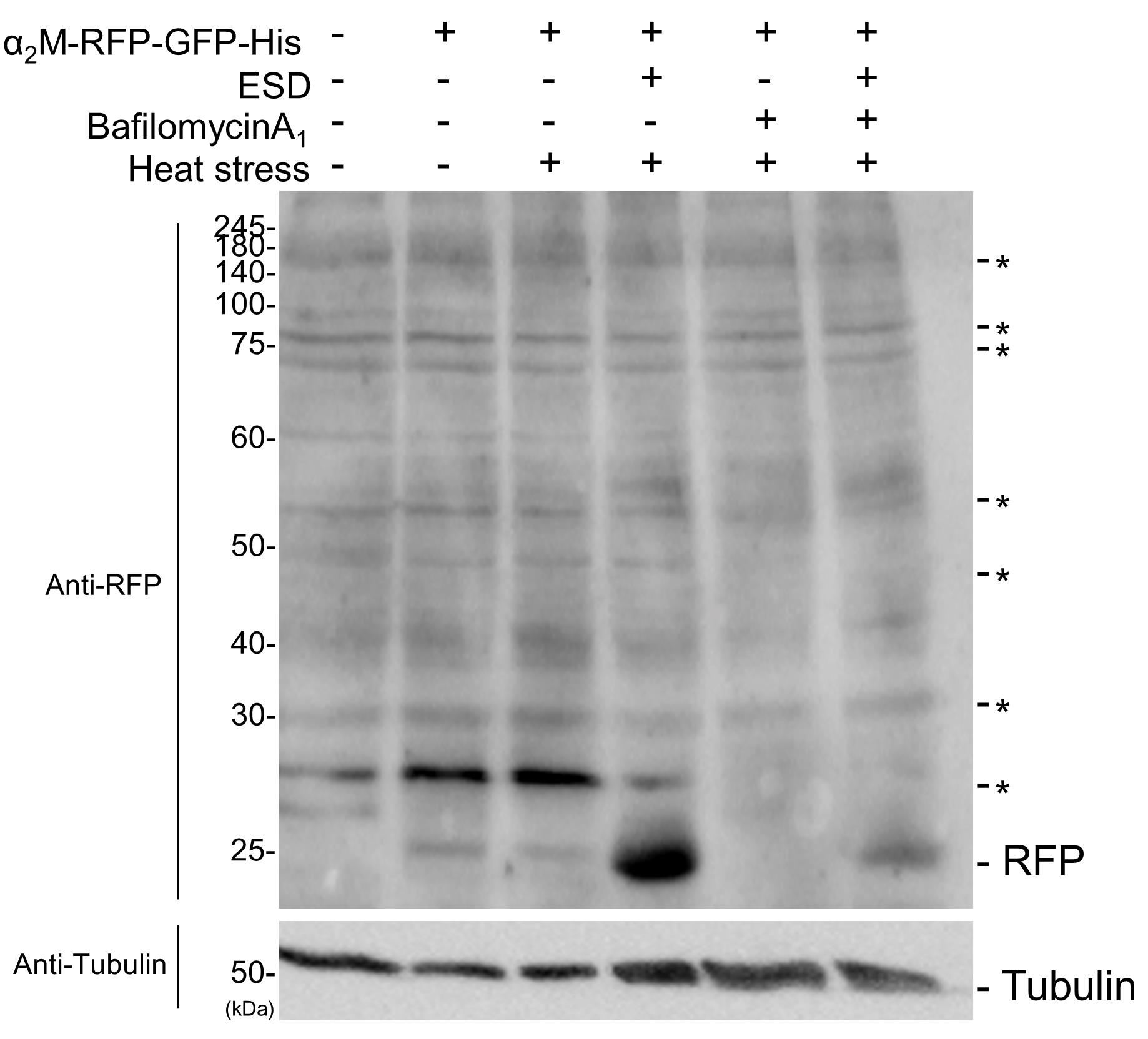
Figure 4. Analysis of α2M-RFP-GFP internalization by immunoblotting. Purified α2M-RFP-GFP-His was added to advanced DMEM/F-12 either alone or together with ESD and preincubated at room temperature or at 50 °C (heat stress) for 1 h. HeLa cells were then cultured in medium with or without bafilomycin A1 for 16 h at 37 °C, followed by analysis by immunoblotting using anti-RFP and tubulin antibodies. Asterisks indicate nonspecific bands.
Validation of protocol
Validation of general applicability to other POIs (Clusterin-RG, Luciferase)
To evaluate the general applicability of this protocol, internalization assays were performed using POIs other than α2M-RFP-GFP-His. Specifically, Clusterin-RFP-GFP-His, an extracellular chaperone, and luciferase, a model misfolded protein, were analyzed for GFP and RFP signals by flow cytometry following internalization. For Clusterin-RFP-GFP-His, an increase in RFP alone was observed, indicating that lysosomal transport and degradation occurred similarly to α2M-RFP-GFP-His (Figure 5). Moreover, when misfolded luciferase was co-administered, the RFP increase was further enhanced, whereas treatment with bafilomycin A1 suppressed the RFP signal. These results are consistent with those observed for α2M, supporting the applicability of this protocol to a broad range of secreted proteins.
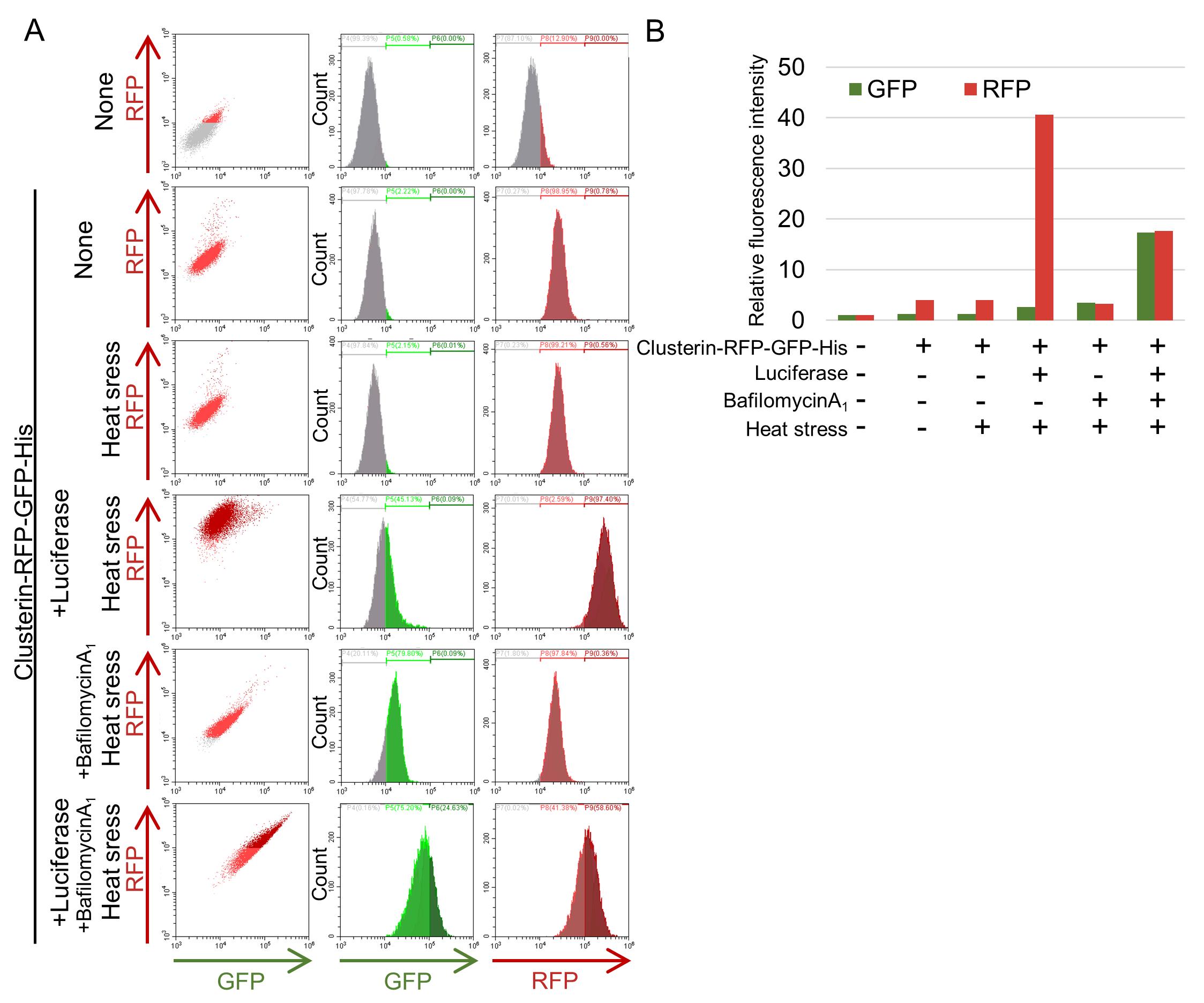
Figure 5. Flow cytometric analysis of Clusterin-RFP-GFP internalization. Purified Clusterin-RFP-GFP-His was added to reduced-serum DMEM/F-12 either alone or together with luciferase and preincubated at room temperature or 42 °C (heat stress) for 1 h. HeLa cells were then cultured in medium with or without bafilomycin A1 for 16 h at 37 °C, followed by analysis using flow cytometry to assess POI internalization. (A) Results from flow cytometric analysis. (B) Graph illustrating the relative fluorescence intensities of GFP and RFP.
General notes and troubleshooting
General notes
1. Advantages and limitations compared with covalent fluorescent labeling: Covalent chemical fluorescent labeling randomly conjugates fluorophores to amino acid residues, which can potentially disrupt the structure or function of the POI. In contrast, this protocol introduces fluorescence by fusing an RFP-GFP tandem tag to either the N- or C-terminus of the POI, ensuring a fixed labeling position and stable fluorescent signals. However, the tandem RFP-GFP tag may still influence the structure or function of the POI. For POIs with unknown structural or epitope information, it is recommended to test both POI-RFP-GFP and RFP-GFP-POI constructs.
2. Distinction between cell surface and endosomal localization: Although flow cytometry enables sensitive detection of changes in fluorescence intensity, it cannot differentiate whether the signal originates from the cell surface or from endosomal/lysosomal compartments. Biochemical analysis using the RFP cleavage assay offers a reliable approach to determine whether the POI-RFP-GFP has undergone lysosomal degradation. By combining flow cytometry with this biochemical approach, whether the POI is internalized and degraded in lysosomes can be accurately evaluated.
3. Immunofluorescence analysis of RFP localization within lysosomes: It is also important to examine the intracellular localization of the RFP signal by immunofluorescence staining using a lysosomal marker (e.g., LAMP1). This approach enables direct confirmation of lysosomal accumulation of the POI-RFP-GFP reporter, thereby complementing the flow cytometric analysis and the immunoblot analysis.
Troubleshooting
1. POI-RFP-GFP is not detected in the culture medium: If the fusion protein is not detectable in the medium, first confirm that the signal peptide is correctly inserted into the plasmid. Next, verify intracellular expression of POI-RFP-GFP by immunoblotting or fluorescence microscopy. If the POI is tissue-specific, secretion may not be observed unless a cell line derived from that tissue type is used.
2. Cells die after 4 days in reduced-serum DMEM/F-12: POI-expressing cells may fail to survive the intended 4-day culture period when maintained in advanced DMEM/F-12. It is essential to avoid nutrient depletion (e.g., medium turning yellow) prior to switching to DMEM/F-12. To maintain cell viability, replenish with fresh growth medium 1 day before exchanging to advanced DMEM/F-12. Alternatively, collect conditioned medium within 3 days to avoid cell death.
3. Low efficiency of POI internalization: Low internalization efficiency may result from insufficient incubation time or suboptimal POI concentration. Internalization is also influenced by cell density; adjusting seeding density may improve uptake. Because internalization capacity varies substantially among cell lines, testing alternative cell lines may also be necessary.
4. Abnormal fluctuations in fluorescence intensity during flow cytometry: Some compounds display strong autofluorescence; therefore, always include a POI-negative control to distinguish true signals from background fluorescence.
5. Lysosomal dysfunction after addition of purified proteins: If lysosomal function appears impaired following the addition of purified proteins, residual imidazole (or any other lysosomotropic components) from the purification process is a likely cause. Lysosomotropic components should be thoroughly removed by ultrafiltration or dialysis before use.
6. Cleaved RFP is not detected in immunoblotting: Because the cleaved RFP fragment may lack portions of its N- or C-terminus, some monoclonal RFP antibodies may not detect it. In such cases, alternative antibodies—such as a polyclonal RFP antibody—should be used to ensure successful detection.
Acknowledgments
This work was supported by JSPS KAKENHI grants (no. 20H03249, 22H04634, 23H04932, and 24K02018 to EI); the JSPS Program for Forming Japan’s Peak Research Universities (J-PEAKS) (grant no. JPJS00420230002 to E.I.); the JST FOREST Program (grant no. JPMJFR204N to E.I.); the Takeda Science Foundation (to E.I.); and the Research Support Program of the Institute for Advanced Academic Research (IAAR) at Chiba University (to E.I.). This protocol was adapted from previous work published in Scientific Reports [9,10].
Competing interests
The authors declare no conflicts of interest.
References
- Mesgarzadeh, J. S., Buxbaum, J. N. and Wiseman, R. L. (2022). Stress-responsive regulation of extracellular proteostasis. J Cell Biol. 221(4): e202112104. https://doi.org/10.1083/jcb.202112104
- Wyatt, A. R., Yerbury, J. J., Ecroyd, H. and Wilson, M. R. (2013). Extracellular Chaperones and Proteostasis. Annu Rev Biochem. 82(1): 295–322. https://doi.org/10.1146/annurev-biochem-072711-163904
- Brown, M. S. and Goldstein, J. L. (1986). A Receptor-Mediated Pathway for Cholesterol Homeostasis. Science. 232(4746): 34–47. https://doi.org/10.1126/science.3513311
- Doherty, G. J. and McMahon, H. T. (2009). Mechanisms of endocytosis. Annu Rev Biochem. 78: 857–902. https://doi.org/10.1146/annurev.biochem.78.081307.110540.
- Chiti, F. and Dobson, C. M. (2017). Protein Misfolding, Amyloid Formation, and Human Disease: A Summary of Progress Over the Last Decade. Annu Rev Biochem. 86(1): 27–68. https://doi.org/10.1146/annurev-biochem-061516-045115
- Tyedmers, J., Mogk, A. and Bukau, B. (2010). Cellular strategies for controlling protein aggregation. Nat Rev Mol Cell Biol. 11(11): 777–788. https://doi.org/10.1038/nrm2993
- Aguzzi, A. and O'Connor, T. (2010). Protein aggregation diseases: pathogenicity and therapeutic perspectives. Nat Rev Drug Discovery. 9(3): 237–248. https://doi.org/10.1038/nrd3050
- Kimura, S., Noda, T. and Yoshimori, T. (2007). Dissection of the Autophagosome Maturation Process by a Novel Reporter Protein, Tandem Fluorescent-Tagged LC3. Autophagy. 3(5): 452–460. https://doi.org/10.4161/auto.4451
- Tomihari, A., Kiyota, M., Matsuura, A. and Itakura, E. (2023). Alpha 2-macroglobulin acts as a clearance factor in the lysosomal degradation of extracellular misfolded proteins. Sci Rep. 13(1):4680. https://doi.org/10.1038/s41598-023-31104-x
- Itakura, E., Chiba, M., Murata, T. and Matsuura, A. (2020). Heparan sulfate is a clearance receptor for aberrant extracellular proteins. J Cell Biol. 219(3): e201911126. https://doi.org/10.1083/jcb.201911126
- Tomihari, A., Chiba, M., Matsuura, A. and Itakura, E. (2021). Protocol for quantification of the lysosomal degradation of extracellular proteins into mammalian cells. STAR Protoc. 2(4): 100975. https://doi.org/10.1016/j.xpro.2021.100975
- Itakura, E., Chen, C. and de Bono, M. (2017). Purification of FLAG-tagged Secreted Proteins from Mammalian Cells. Bio Protoc. 7(15): e2430. https://doi.org/10.21769/bioprotoc.2430
- Katayama, H., Kogure, T., Mizushima, N., Yoshimori, T. and Miyawaki, A. (2011). A Sensitive and Quantitative Technique for Detecting Autophagic Events Based on Lysosomal Delivery. Chem Biol. 18(8): 1042–1052. https://doi.org/10.1016/j.chembiol.2011.05.013
- Costantini, L. and Snapp, E. (2013). Probing Endoplasmic Reticulum Dynamics using Fluorescence Imaging and Photobleaching Techniques. Curr Protoc Cell Biol. 60(1): ecb2107s60. https://doi.org/10.1002/0471143030.cb2107s60
- Gibson, D. G. (2011). Enzymatic Assembly of Overlapping DNA Fragments. Meth Enzymol. 498: 349–361. https://doi.org/10.1016/b978-0-12-385120-8.00015-2
- Liu, Z., Wang, Y., Zhao, S., Zhang, J., Wu, Y. and Zeng, S. (2014). Imidazole inhibits autophagy flux by blocking autophagic degradation and triggers apoptosis via increasing FoxO3a-Bim expression. Int J Oncol. 46(2): 721–731. https://doi.org/10.3892/ijo.2014.2771
- Brunelle, J. L. and Green, R. (2014). Coomassie Blue Staining. Meth Enzymol. 541: 161–167. https://doi.org/10.1016/b978-0-12-420119-4.00013-6
Article Information
Publication history
Received: Dec 10, 2025
Accepted: Jan 28, 2026
Available online: Feb 6, 2026
Published: Mar 5, 2026
Copyright
© 2026 The Author(s); This is an open access article under the CC BY-NC license (https://creativecommons.org/licenses/by-nc/4.0/).
How to cite
Bun, S., Kamikawa, K., Matsuura, A. and Itakura, E. (2026). Quantifying Lysosomal Degradation of Extracellular Proteins With a Fluorescent Protein-Based Internalization Assay. Bio-protocol 16(5): e5619. DOI: 10.21769/BioProtoc.5619.
Category
Cell Biology > Cell-based analysis > Endocytosis
Biochemistry > Protein > Degradation
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.