- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Multiplexing Thermotaxis Behavior Measurement in Caenorhabditis elegans
(*contributed equally to this work) Published: Vol 12, Iss 7, Apr 5, 2022 DOI: 10.21769/BioProtoc.4370 Views: 2929
Reviewed by: Manish ChamoliKyung Suk LeeYusuke TominaAnonymous reviewer(s)
Original research article
The authors used this protocol in:

Mar 2021
Advertisement



Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Related protocols
Abstract
Thermotaxis behaviors in C. elegans exhibit experience-dependent plasticity of thermal preference memory. This behavior can be assayed either at population level, on linear temperature gradients, or at the individual animal level, by radial isothermal or microfluidic tracking of orientation. These behaviors are low-throughput as well as variable, due to the inherent sensitivity to environmental perturbations. To facilitate reproducible studies, we describe an updated apparatus design that enables simultaneous runs of three thermal preference assays, instead of single-run assays described previously. By enabling parallel runs of control and experimental conditions, this set-up enables more throughput and rigorous assessment of behavioral variability.
Keywords: C. elegansBackground
The C. elegans nervous system comprises 56 glia or glia-like cells and 300 neurons. This simple nervous system, mapped in both sexes, drives various sensory and reproductive behaviors. The invariant cell-lineages and cell-cell contacts, mapped connectome, and powerful genetics of this setting facilitate the speedy and precise molecular-genetic dissection of the circuit biology driving these behaviors (Singhvi and Shaham, 2019). One such behavior is thermotaxis, or temperature-dependent sensory behavior. Thermotaxis is not an innately hard-wired behavior within physiological temperature range; rather, the animal’s thermotaxis memory and preference exhibits experience-dependent plasticity on a timescale of hours, based on the animal’s cultivation conditions (Mori and Ohshima 1995; Ramot et al., 2008; Goodman et al., 2014; Aoki and Mori 2015; Goodman and Sengupta, 2019).
C. elegans thermotactic behaviors were first described in 1975, and subsequently dissected by many labs (Hedgecock and Russell, 1975; Garrity et al., 2010; Sengupta and Garrity; 2013; Goodman et al., 2014; Goodman and Sengupta, 2019). Briefly, when C. elegans is raised at a certain temperature (Tc = cultivation temperature), it will migrate up or down a temperature gradient toward Tc. Movement toward warmer temperatures is noted as the animal’s thermophilic drive (positive thermotaxis), while movement down a gradient is the cryophilic drive (negative thermotaxis). At the preferred Tc, animals track isothermal contours (Hedgecock and Russell, 1975; Garrity et al., 2010; Goodman et al., 2014). Hedgecock and Russell (1975) demonstrated C. elegans thermotactic behavior in two assay conditions, a linear thermal gradient of 0.5°C/cm (chosen for its similarity with gradients in moist soil, the animal’s natural habitat), and isothermal tracks of individual animals in a radial gradient. For radial gradients, Hedgecock and Russell (1975) placed a vial of frozen glacial acetic acid (melting point 16.6°C) on the bottom-center of a plastic agar plate. For linear gradients, an aluminum cube was bolted on either end of a thin (1.3 cm) aluminum slab, and each cube “leg” was immersed into a thermostatically regulated water bath (5°C or 35°C on either end). Four Petri plates were placed in line along this slab (with the center at ~20°C), such that each equilibrated to a different temperature, from 12.9–27.1°C. Subsequent studies utilized Peltier-effect thermoelectric elements and resistive cartridge heaters to generate analogous stable linear temperature gradients on single agar plates (Ryu and Samuel 2002; Goodman et al., 2014).
However, while many labs were able to replicate these assay results in the years to follow, some groups obtained divergent results (Kimata et al., 2012). Careful analyses by Jurado et al. (2010) indicated that the assay variability across laboratories possibly reflected differences in the temperature gradients of the assay chambers. Specifically, gradients of 1°C/cm or less elicited stable thermophilic behaviors and steeper gradients suppressed them (Ramot et al. 2008; Jurado et al., 2010; Kimata et al., 2012).Jurado et al. (2010) posited that this approximated the environmental temperature gradients in soil that animals would experience (Ramot et al., 2008; Jurado et al., 2010). It also matched with computational modeling of animal behavior (Nakazato and Mochizuki, 2009). Furthermore, other published and unpublished observations indicated that thermotaxis is an unusually sensitive behavior, which is readily altered by conditions like humidity (and by extension, seasonal variation), starvation, population density, and time-frame of the assay runs. Together, these suggest that careful monitoring of animal growth and assay conditions are imperative for definitive assessment of these behaviors.
The AFD sensory neuron is the predominant driver of animal thermosensory behaviors (Mori and Ohshima, 1995). Its neuron receptive-ending (AFD-NRE) is a microvilli organelle with a single cilium, embedded entirely within the sense-organ amphid sheath glia (AMsh). We and others have shown that AMsh glia impact the animal’s thermosensory behaviors and memory, by regulating AFD-NRE shape. AMsh glia deploy multiple mechanisms to do so, including regulating the ionic milieu around AFD-NRE, and engulfing AFD-NRE fragments (Singhvi et al., 2016; Yoshida et al., 2016; Raiders et al., 2021).
The thermotaxis assay protocol described here (used in Raiders et al., 2021) is as described previously (Goodman et al., 2014), with a modified apparatus design that enables tandem runs of three assays i.e., control and experimental strains simultaneously. This modification provides two advantages: (i) it allows easy tracking and assessment of the inherent behavioral variability of thermotaxis assays, and (ii) it increases the throughput of these time-consuming assays. This set-up also enables automated video-tracking of animal locomotory behaviors within the assay chamber.
Environmental Room
All behavior assays are conducted in an environmentally controlled room set at 20 ± 0.5°C and 32 ± 5% relative humidity. Should such a room not be accessible, it is strongly recommended that (a) control groups are run interspersed with the experimental groups; (b) humidity is measured by a hygrometer in both growth and assay areas, (c) temperature is monitored in incubator and assay areas, and (d) dates are tracked, to monitor for potential ambient seasonal variability.
Materials and Reagents
Worm pick
5¾ Disposable Pasteur Pipets (Thermo Fisher Scientific, Fisherbrand, catalog number: 13-678-20B)
99.95% Pt Wire, 30 gauge (Genesee Scientific, catalog number: 59-1M30P)
C. elegans strains
N2 (wild type strain) (Caenorhabditid Genetics Center (CGC), catalog number: N2)
Bacteria Strains for nematode growth
OP-50 (Caenorhabditid Genetics Center (CGC), catalog number: OP-50)
Luria Broth (for growing OP-50 bacteria)
Luria Broth 1 L Buffered Capsules [Miller's LB Broth] (Research Products International, catalog number: L24045-1000.0)
NGM agar plates for growth and assay (see Recipes)
Ingredients:
Bacto agar (Becton Dickinson and Company, BD, catalog number: 214010, lot: 1152216)
Bacto Peptone (Thermo Fisher Scientific, Gibco, catalog number: 211677, lot: 01896889)
Magnesium Sulfate Heptahydrate (Thermo Fisher Scientific, Fisher Bioreagents, catalog number: BP213-1, lot: 175995)
Potassium Phosphate Monobasic (Thermo Fisher Scientific, Fisher Bioreagents, catalog number: BP362-500, lot: 204665)
Potassium Phosphate Monobasic (Thermo Fisher Scientific, Fisher Bioreagents, catalog number: BP363-500, lot: 197723)
Calcium Chloride Dihydrate (Thermo Fisher Scientific, Fisher Chemical, catalog number: C70-500, lot: 187321)
Choloesterol (Sigma-Aldrich, Sigma, catalog number: C8667-100G, lot: SLCG0969)
Supplies:
Drosophila Supplies Petri Dishes (Thermo Fisher Scientific, Fisherbrand, catalog number: AS4050) (for standard nematode growth)
Square Petri Dish with Grid (Simport Scientific, catalog number: D210-16) (for assay)
M9 buffer (see Recipes)
Sodium Phosphate Dibasic Anhydrous (Thermo Fisher Scientific, Fisher Chemical, catalog number: S374-500)
Potassium Phosphate Monobasic (Thermo Fisher Scientific, Fisher Bioreagents, catalog number: BP363-500, lot: 197723)
Sodium Chloride (Thermo Fisher Scientific, Fisher Chemical, catalog number: S271-500)
Ammonium Chloride (Thermo Fisher Scientific, Fisher Chemical, catalog number: A661-500)
Bleach Solution
Sodium hypochlorite solution (Sigma-Aldrich, catalog number: 42050-250 ml)
Sodium hydroxide (Thermo Fisher Scientific, Fisher Chemical, catalog number: S318-500)
M9 (see above)
15 mL Falcon Conical Centrifuge Tubes (Thermo Fisher Scientific, Fisher scientific, catalog number: 05-527-90)
S-Basal buffer (see Recipes)
Sodium Chloride (Thermo Fisher Scientific, Fisher Chemical, catalog number: S271-500)
Potassium Phosphate Monobasic (Thermo Fisher Scientific, Fisher Bioreagents, catalog number: BP362-500)
Potassium Phosphate Monobasic (Thermo Fisher Scientific, Fisher Bioreagents, catalog number: BP363-500)
Choloesterol (Sigma-Aldrich, Sigma, catalog number: C8667-100G)
Microcentrifuge tubes (Lab Force, Posi-Click, catalog number: 1149K01)
Kim wipes (Kimberly-Clark Professional, catalog number: 34155)
Glycerol (Sigma-Aldrich, Sigma Life Sciences, catalog number: G5516-500ML)
Lasergrip 1080 Laser Thermometer (Etekcity)
Chloroform (Thermo Fisher Scientific, Fisher Chemical, catalog number: C289-500)
Equipment
A schematic of the setup is presented, with explanations in Figure 1. The design, which creates a stable thermal gradient in one direction by using thermoelectric coolers, is similar to earlier work in a number of model systems, including C. elegans. The unique feature here is that it holds three bio-assay dishes for parallel experiments. The controlled thermal gradient platform was constructed with the parts listed below, along with three custom aluminum blocks, which can be machined using the drawings included in Figure 2. Assembly instructions are included, along with additional images in Figure 3. Numbers in parentheses following a part name indicates the number of each part needed.
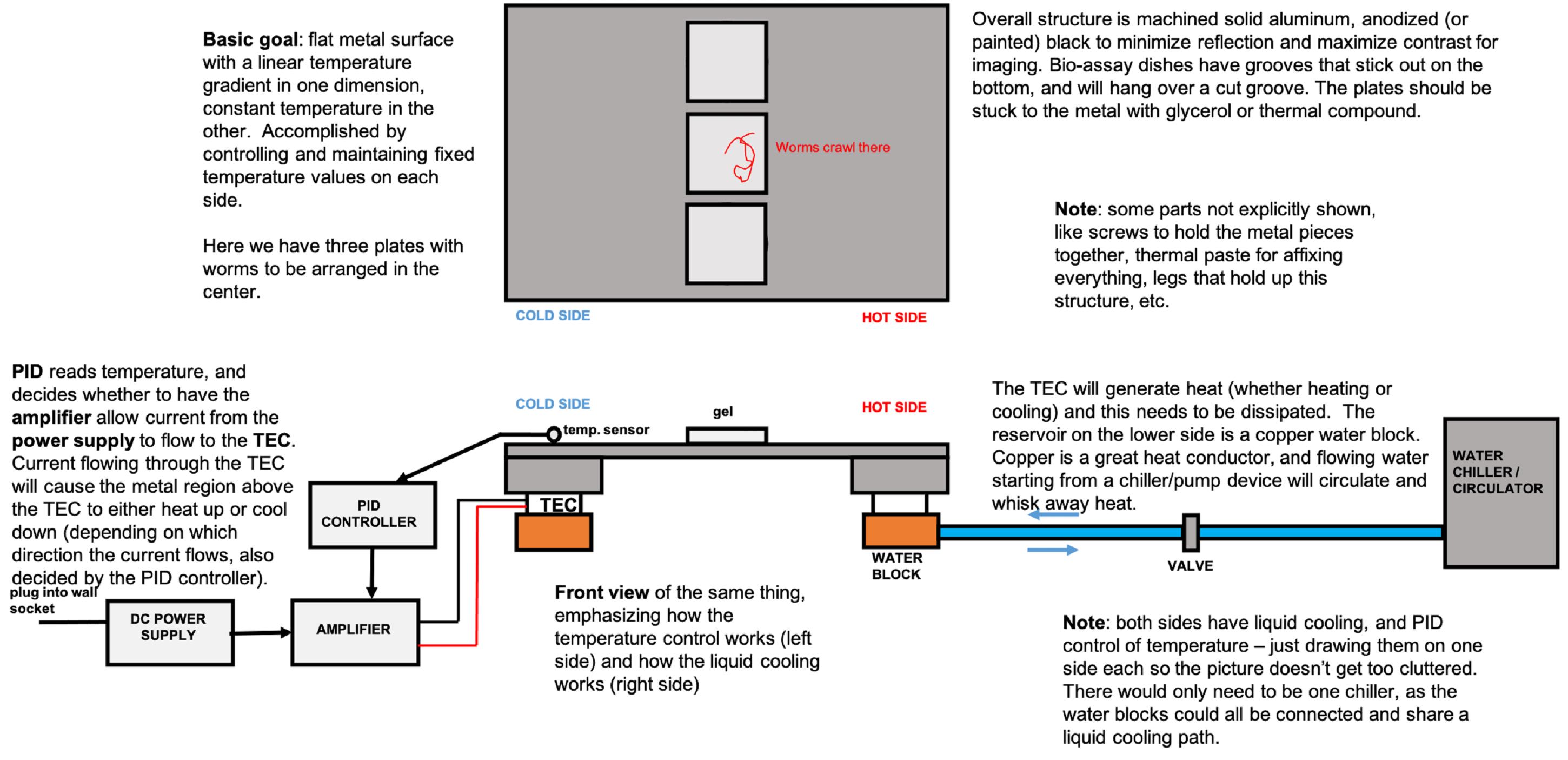
Figure 1. Schematic of the structure of the linear thermal gradient device, with notes explaining the function of some parts. Basic Goal: flat metal surface with a linear temperature gradient in one dimension, and constant temperature in the other. Accomplished by controlling and maintaining fixed temperature values on each side. Overall structure is machined solid aluminum, anodized (or painted) black, to minimize reflection and maximize contrast for imaging. Bio-assay dishes have grooves that stick out on the bottom, and will hang over a cut groove. The plates should be stuck to the metal with glycerol or thermal compound. Note: Some parts not explicitly shown like screws to hold the metal pieces together, thermal paste for affixing everything, legs that hold up this structure, etc. Here, we have three plates with worms to be arranged in the center. PID reads temperature and decides whether to have the amplifier allow current from the power supply to flow to the TEC. Current flowing through the TEC will cause the metal region above the TEC to either heat up or cool down (depending on which direction the current flows, and also decided by the PID controller). Front view of the same thing, emphasizing how the temperature control works (left side) and how the liquid cooling works (right side). The TEC will generate heat (whether heating or cooling) and this needs to be dissipated. The reservoir on the lower side is a copper water block. Copper is a great heat conductor, and flowing water starting from a chiller/pump device will circulate and whisk away heat. Note: Both sides have liquid cooling and PID control of temperature – here just drawn on one side each, so the picture doesn’t get too cluttered. There would only need to be one chiller, as the water blocks could all be connected, and share a liquid cooling path.

Figure 2. Machine drawings for the pieces for the linear thermal gradient device. LEFT: horizontal gradient plate. RIGHT: reservoir blocks (make 2). The material is aluminum (6061). Dimensions (red) are in inches. The four holes on the underside of the reservoir blocks (the D&T 6–32 labels) may need to be different, depending on the water block chosen. The horizontal tapped holes in the sides of the reservoir blocks are not required, but might be convenient for attaching a structure to hold the (optional) camera.

Figure 3. Photographs of the assembled thermal gradient setup. TOP: Assembled electronics for one side of the setup. The three main components are (1) power supply (silver colored box), (2) PID controller (black box), and (3) H-bridge amplifier (exposed circuit piece). The white/red braided wire on the right is the temperature sensor and should adhere to the top of the horizontal gradient plate (on one side, above the reservoir block), and the red/black wire should connect/solder to both TECs on one side of the horizontal gradient plate, and the TECs should be connected in parallel. BOTTOM: Assembled version with electronics on both sides and the crawling arena, with room for three square bio-assay dishes, in the center. (a) PID controllers; (b) bio-assay dish locations; (c) temperature sensors; (d) reservoir blocks; (e) legs/feet; (f) quick-disconnect valve; (g) liquid cooling tubing.
Metal plate structure:
Horizontal gradient plate, a 12” × 12” × 1/4” aluminum block (McMaster-Carr, catalog number: 8975K142)
Reservoir blocks (2), 12” × 4” × 1.5” aluminum blocks (MMcMaster-Carr, catalog number: 8975K257)
Legs to hold structure (McMaster-Carr, catalog number: 93330A652)
Feet for damping vibrations (McMaster-Carr, catalog number: 9995T91)
Heat sink compound (McMaster-Carr, catalog number: 10405K83)
Temperature control electronics
Temperature sensors, 100 ohm RTDs (2) (McMaster-Carr, catalog number: 6568T43)
PID controllers (2) (Accuthermo Technology, catalog number: FTC100D)
H-bridge amplifiers (2) (Accuthermo Technology, catalog number: FTX700D)
Power supplies (2), 12 V, 450 W (Digi-Key, catalog number: 1866-4473-ND)
Zip cord 16 awg, 15 A, 50 ft spool (McMaster-Carr, catalog number: 7870T31)
Ribbon cable, 26 awg, 50 ft (Digi-Key, catalog number: MC010M-50-ND)
Thermoelectric coolers (TECs), 47 W, RTV-sealed (Custom Thermoelectric, catalog number: 12711-5L31-05CQ)
Liquid cooling / heat dissipation
Water blocks, copper (4) (Custom Thermoelectric, catalog number: WBA-1.62-0.55-CU-01)
PVC tubing, ½" ID, 25 ft (McMaster-Carr, catalog number: 5231K228)
Quick disconnect valve, plug (4) (McMaster-Carr, catalog number: 5385K25)
Quick disconnect valve, socket (4) (McMaster-Carr, catalog number: 5385K55)
Circulating chiller (VWR, catalog number: 89202-966)
Lighting and image acquisition (optional)
LED strips, red, kit (Inspired LED, model: super bright flexible strip kit – red: #3739)
Camera, 5 MP, USB3.0 (Mightex Systems, catalog number: SME-B050-U)
Lens for camera, 8 mm (Edmund Optics, catalog number: 56-786)
Metal plate structure.
Machine aluminum blocks as diagrammed in Figures 1 and 2. Clean surfaces with a paper cleaning wipe using water and then 70% ethanol. Then, apply a thin layer of heat sink compound to the top of each reservoir block, place the horizontal gradient plate atop the reservoir blocks, and secure with screws. Secure legs and feet with set screws.
Temperature control electronics
Adhere a temperature sensor to the top of the horizontal gradient plate, directly above the center of each reservoir block. Apply a thin layer of heat sink compound to both sides of all four TECs, and place two on the underside of each reservoir block, then attach a copper water block (with screws into the reservoir block) to each one to hold in place. Connect each temperature sensor to a PID controller, then connect each PID controller to an H-bridge amplifier (four wires using ribbon cable). Connect each 12 V power supply to the input of each H-bridge amplifier (using zip cord). Connect the output of each H-bridge amplifier (also zip cord) to a pair of TECs (in parallel) on one of the reservoir blocks.
Temperature control liquid cooling
Using PVC tubing, connect the circulating chiller output to the water blocks, going through each of the four in series, then back to the circulating chiller input. Quick-disconnect can be used between the chiller and the thermal gradient device. When running the chiller, typically set to a low level, but above freezing.
Lighting and image acquisition (optional).
Cut and shape LED strips as desired, forming a rectangle, so that the LEDs point inward toward the crawling arena from the sides (dark field illumination). Position the camera (with lens) directly above the arena in a bird’s eye view formation. See Figure 4.
To operate the linear spatial gradient, circulate cold water through the water blocks with the chiller, then turn on electronics. Set the PID controller on each side to desired temperature. If needed, the PID constants can be set using the auto-tune feature. Once both temperatures reach their set points, a smooth and stable linear gradient should be established across the horizontal plate.

Figure 4. Photographs showing optional lighting and camera placements. LEFT: Aluminum frame made from construction rails (dashed yellow lines for emphasis), which holds a CCD camera looking down at the arena. RIGHT: LED bars pointed inwards towards a square arena (dashed red lines for emphasis) for dark field illumination. Note that this photograph is from a different setup, but the placement would work the same way.
Procedure
Assay plate preparation
NGM agar plates for growth and assay (see Recipes)
Prepare plates less than 1 week before assay. Keep plates sealed at 4°C before use.
Prepare standard NGM plate media as per established protocol. Cool to 50°C.
Note: We usually make about 20 plates per week when running assays. To pour this many plates, make 250 mL of media.
Pipette 12 mL of media into 20 individual 10-cm2 Petri dishes.
Note: It is important that there are no bubbles in the plate, as worms will burrow into these.
Let cool for 3 h, and then store in a sealed plastic bag at 4°C.
Use assay plates that are 2–5 days old. In our hands, both too-fresh and too-old plates impact assays.
Worm preparation
M9 buffer (see Recipes)
Grow 3–4 plates up, containing gravid adult worms of each genotype/condition.
Bleach these plates according to standard protocol in a 15-mL Falcon tube, to obtain clean eggs (Stiernagle, 2006).
After the final wash, suspend eggs in 5 mL of 1× M9 and rotate overnight, to obtain synchronized L1 larvae.
The next day, set tube upright ~30 min so L1s sink to the bottom of the M9, or spin them down at 300 × g for 1 min.
Remove excess M9 with a glass pipette.
Use a glass pipette to move worms from the tube onto a seeded NGM plate.
Note: It is important to have enough worms to perform the assay (~200 worms per replicate). It is also important not to crowd the plate or starve the worms. Any amount of starvation after synchronization can alter the outcome of the assay.
Grow the worms at the cultivation temperature being analyzed (typically 15°C or 25°C) until the worms have reached young adulthood (Table 1).
Table 1. Approximate growth times for N2 from synchronized L1s to adulthood (Stiernagle, 2006)
Cultivation Temperature Time from L1 to young adult 15°C ~96 h 20°C ~60 h 25°C ~48 h
Note: This is usually the day after the worms are L4 larvae. The amount of time needed to get the worms to adulthood will vary with temperature. It is important to raise all control and experiment worms in parallel, so that they experience the same conditions and are at the same age when conducting the assay.
Pre-assay preparation
One hour before beginning assay, remove assay plates from 4°C, and allow them to warm up to room temperature.
Turn on the water pump and thermo-regulators. Before starting the assay, ensure that water is flowing, and that each side of the thermotaxis rig is at the correct temperature.
The gradient for the assay is 1°C/cm, with the very center of the plate being 20°C. Adjustment of each thermo-regulator may be necessary to have this gradient. This results in the left most edge of the plate being at ~16.5°C, and the rightmost being at ~23.5°C.
Performing the assay
S-Basal buffer (see Recipes)
Note: The next steps should be carried out as quickly as possible before the worms have time to adjust to room temperature. We typically get the worms from cultivation temperature to the thermotaxis rig in ~5–10 min (Steps 1–3 shown in Video 1; Steps 4–7 shown in Video 2).
 Video 1. Placing worms on assay plate.
Video 1. Placing worms on assay plate.Remove the first plate of worms. Using a Pasteur pipette, place ~2 mL of room temperature S-basal on the plate and lightly shake, to allow worms to detach from the bacterial surface.
Tilt the plate and pick up as much of the worm slurry as possible. Pipette into a microcentrifuge tube.
Either allow the worms to settle to the bottom of the tube or lightly spin them down with a tabletop microcentrifuge for 3 s.
Note: Depending on the centrifuge, spinning down the worms may damage them. If this is a concern, we recommend opting for letting the worms settle to the bottom of the plate.
Gently pipette as much S-Basal off the worms as possible. The goal is to remove the maximum possible number of bacteria.
Dispose of the S-Basal and gently fill the tube with clean S-Basal.
Gently mix the worms and then spin down or allow them to settle.
Pick up the worms in the pipette with minimal S-Basal.
Hold the pipette upright for a few moments, to allow the worms to settle to the bottom.
Pipette the worms (<200 worms) onto the center of the assay plate.
Note: Worm density is critical. Both too sparse and too dense inversely impact assay fidelity. Further, too low and there won’t be enough worms to perform the assay. Too high will lead to the worms clumping together at the center of the plate.
Take a Kim wipe and fold it in half along the crease. Take one of the creased corners and twist it to make a long point.
Under the dissecting microscope carefully soak up the S-Basal from the center of the plate via the capillary action from the tip of the Kim wipe.
Note: It is important to try and avoid the worms. They can be pulled into the Kim wipe or damaged enough to impair proper locomotion on the assay plate. We find that quick dabs around different edge-points of the drop works best.
Soak up S-Basal until all worms are freely crawling out of the drop-site on the plate.
Note: Do not pull up too much liquid, as this will dry and tear the surface of the plate. Worms can burrow into the plate through these micro-abrasions, complicating the assay.
Using a pipette, transfer 5 mL of glycerol to the center of the back plate of the thermotaxis rig.
Carefully lay one edge of the assay plate on the corresponding edge of the back plate. Slowly lay the assay plate onto the glycerol, giving it time to spread evenly and without bubbles on the bottom of the assay plate. Else, the air pockets can impede uniform gradient heat transfer.
The plate should float on glycerol evenly along its entire bottom surface, including all edges.
When the plate is all the way down, ensure that it is straight and properly fitted to the back plate.
Begin a timer for 45 min.
Tandem assay set-up
We run three trials of each condition on separate days. Each day, we include a wild type control and up to two test conditions.
On each day, we set up each position in descending order from 1 to 3 (see Table 1). Each position is set up 10 min after the previous position, they are run in tandem for some time, and each plate removed after it has been on the rig for 45 min.
To address variability between positions on the rig, for each trial we rotate each condition through each position on the rig (see Table 2).
Table 2. Scheme for rotating each condition on the thermotaxis rig
Position Trial 1 Trial 2 Trial 3 Position 1 Wild type Test 2 Test 1 Position 2 Test 1 Wild type Test 2 Position 3 Test 2 Test 1 Wild type
Scoring the assay
At the end of the 45 min, apply 1 mL of chloroform to the bottom of assay plate lid, invert the assay plate over it, and remove from the rig (Video 2). This will immobilize the worms for counting.
When finished with the assay, clean the back of the plate with paper towels and 70% ethanol to remove residual glycerol.
Video 2. Placing assay plate on thermotaxis rig.
Data analysis
The assay plate is a 6 × 6 grid (Figure 5):
Make a template like the grid above. For each plate, count the number of worms in each square.
Note: We count worms that are on the edges of the plate but still on the agar as being within the nearest bin.
Calculate the percentage of worms that are in each column (1–6) along the gradient.
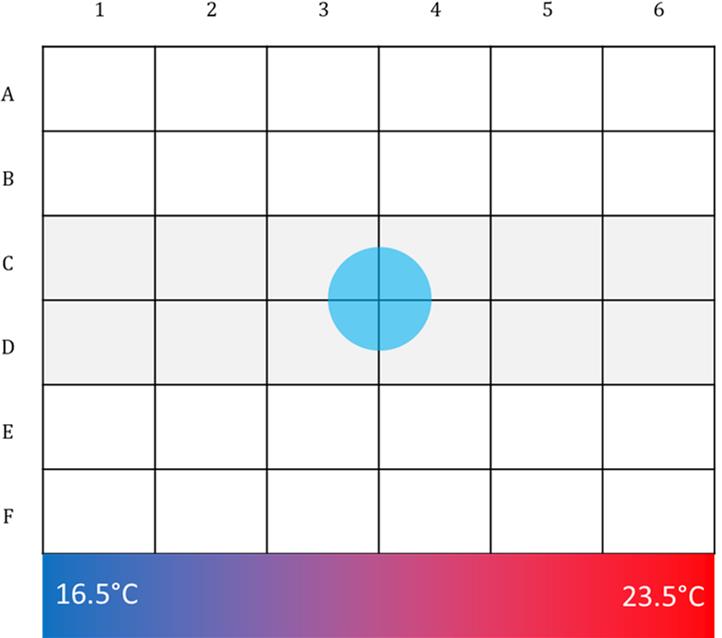
Figure 5. Scoring Grid for Thermotaxis Assay.Run a minimum of three trials for each genotype/condition on at least three separate days.
Censor all runs on days when there are obvious issues with worm locomotion, or the control/wild type worms do not behave as expected.
Make sure to cycle each condition through each position of the thermotaxis rig, to address potential variation (Table 2). At least three consistent runs are required for data analyses.
Record the number of animals in each square following the experiment.
Note: If you do not wish to immediately score the animals, you may take a picture of the whole plate immediately following paralysis of the worms.
Censor the middle two rows (C and D). We prefer this conservative approach to avoid the confound of dead/damaged worms in the center. Pool worms into 6 bins, representing each column.
Combine the number of worms in each bin and calculate the proportion of total worms per bin.
These data can be graphed and compared to controls/wild type qualitatively.
The thermotaxis index (as described in Goodman et al., 2014) can be used to quantitatively compare between conditions.
Notes
| Problem | Possible causes and solutions |
| Worms form clumps | Reasons can include: 1. Density: Worms will clump if their density hits a certain threshold. Be sure to plate ~200 worms per assay. If this continues to be an issue, spread the worms out more in the drop before wiping up the liquid with the Kimwipe. 2. Liquid: Be sure to wipe up the liquid well. 3. We find that humidity also affects clumping. To avoid this, our assays are performed in a temperature- and humidity-controlled room. Worms to be tested are also grown in an incubator in this room. In our hands, this yields greater consistency in the assays across days. |
| Control worms move to the wrong side | Reasons for this can include: 1. Worms were starved recently before the test. 2. The gradient is incorrect. Check the gradient is correct by measuring the left and right sides of the plate before performing the assay. Note: It is helpful to regularly monitor the temperature of the cultivation incubator (EasyLog EL-SIE-1 Temp Logger) to ensure it is accurate and consistent. |
| Worms burrow into plates | 1. Be sure not to score or mark the plate. 2. Check that the plate has not dried or cracked. |
| Worms do not move from middle of plate | 1. This will occur if worms are damaged or left in liquid for too long. |
| Other reasons for inconsistent results | 1. Either culture or assay plates are too wet/dry. Local plate humidity also affects assays. 2. Air vent draft around the assay plate. 3. Uneven stimuli around assay plate such as perfume or other smells, light, or sound. 4. Genetic drift. Thaw fresh worm stocks every ~3 months and use outcrossed strains (4×–8×). 5. Contamination on plates. |
Recipes
Recipes for NGM-agar, M9 buffer, and S-Basal buffer were standard formulations, as described in WormBook: (http://www.wormbook.org/chapters/www_strainmaintain/strainmaintain.html).
Acknowledgments
This work was supported by National Institute of Health F31NS118958-01A1 and T32T32AG066574 to SR (AS laboratory); Glenn Foundation for Medical Research and AFAR Junior Faculty Grant, Simons Foundation/SFARI Grant 488574, National Institute of Health R01 NS114222, and Anderson Foundation and the Marco J. Heidner Foundation Pilot Fund to A.S. Data from this apparatus is described in Raiders et al., eLife 2021 (doi: 10.7554/eLife.63532). We sincerely thank Maria Purice for help with filming the videos.
Competing interests
The authors declare no competing interests.
Ethics
Per NIH guidelines, this study does not require approval from ethical oversight committees.
References
- Aoki, I. and Mori, I. (2015). Molecular biology of thermosensory transduction in C. elegans. Curr Opin Neurobiol 34: 117-124.
- Garrity, P. A., Goodman, M. B., Samuel, A. D. and Sengupta, P. (2010). Running hot and cold: behavioral strategies, neural circuits, and the molecular machinery for thermotaxis in C. elegans and Drosophila. Genes Dev 24(21): 2365-2382.
- Goodman, M. B., Klein, M., Lasse, S., Luo, L., Mori, I., Samuel, A., Sengupta, P. and Wang, D. (2014). Thermotaxis navigation behavior. WormBook: 1-10.
- Goodman, M. B. and P. Sengupta (2019). How Caenorhabditis elegans Senses Mechanical Stress, Temperature, and Other Physical Stimuli. Genetics 212(1): 25-51.
- Hedgecock, E. M. and Russell, R. L. (1975). Normal and mutant thermotaxis in the nematode Caenorhabditis elegans. Proc Natl Acad Sci U S A 72(10): 4061-4065.
- Jurado, P., Kodama, E., Tanizawa, Y. and Mori, I. (2010). Distinct thermal migration behaviors in response to different thermal gradients in Caenorhabditis elegans. Genes Brain Behav 9(1): 120-127.
- Kimata, T., Sasakura, H., Ohnishi, N., Nishio, N. and Mori, I. (2012). Thermotaxis of C. elegans as a model for temperature perception, neural information processing and neural plasticity. Worm1(1): 31-41.
- Mori, I. and Ohshima, Y. (1995). Neural regulation of thermotaxis in Caenorhabditis elegans. Nature 376(6538): 344-348.
- Nakazato, K. and A. Mochizuki (2009). Steepness of thermal gradient is essential to obtain a unified view of thermotaxis in C. elegans. J Theor Biol 260(1): 56-65.
- Raiders, S., Black, E. C., Bae, A., MacFarlane, S., Klein, M., Shaham, S. and Singhvi, A. (2021). Glia actively sculpt sensory neurons by controlled phagocytosis to tune animal behavior. Elife 10: e63532.
- Ramot, D., MacInnis, B. L., Lee, H. C. and Goodman,. M. B. (2008). Thermotaxis is a robust mechanism for thermoregulation in Caenorhabditis elegans nematodes. J Neurosci 28(47): 12546-57.
- Ryu, W. S. and A. D. T. Samuel (2002). Thermotaxis in Caenorhabditis elegans Analyzed by Measuring Responses to Defined Thermal Stimuli. J Neurosci 22(13): 5727-5733.
- Sengupta, P. and Garrity, P. (2013). Sensing temperature. Curr Biol 23(8): R304-307.
- Singhvi, A., Liu, B., Friedman, C. J., Fong, J., Lu, Y., Huang, X. Y. and Shaham, S. (2016). A Glial K/Cl Transporter Controls Neuronal Receptive Ending Shape by Chloride Inhibition of an rGC. Cell 165(4): 936-948.
- Singhvi, A. and Shaham, S. (2019). Glia-Neuron Interactions in Caenorhabditis elegans. Annu Rev Neurosci 42: 149-168.
- Stiernagle, T. (2006). Maintenance of C. elegans. WormBook: 1-11.
- Yoshida, A., Nakano, S., Suzuki, T., Ihara, K., Higashiyama, T. and Mori, I. (2016). A glial K+/Cl- cotransporter modifies temperature-evoked dynamics in Caenorhabditis elegans sensory neurons. Genes Brain Behav 15(4): 429-40.
Article Information
Copyright
![]() Raiders et al. This article is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0).
Raiders et al. This article is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0).
How to cite
Readers should cite both the Bio-protocol article and the original research article where this protocol was used:
- Raiders, S., Klein, M. and Singhvi, A. (2022). Multiplexing Thermotaxis Behavior Measurement in Caenorhabditis elegans. Bio-protocol 12(7): e4370. DOI: 10.21769/BioProtoc.4370.
- Raiders, S., Black, E. C., Bae, A., MacFarlane, S., Klein, M., Shaham, S. and Singhvi, A. (2021). Glia actively sculpt sensory neurons by controlled phagocytosis to tune animal behavior. Elife 10: e63532.
Category
Neuroscience > Sensory and motor systems > Animal model
Neuroscience > Basic technology
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.