- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Simple Scalable Protein Expression and Extraction Using Two-stage Autoinducible Cell Autolysis and DNA/RNA Autohydrolysis in Escherichia coli
Published: Vol 12, Iss 2, Jan 20, 2022 DOI: 10.21769/BioProtoc.4297 Views: 3551
Reviewed by: Anonymous reviewer(s)
Original research article
The authors used this protocol in:

Sep 2020
Advertisement


Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Abstract
Recombinant protein expression is extensively used in biological research. Despite this, current protein expression and extraction methods are not readily scalable or amenable for high-throughput applications. Optimization of protein expression conditions using traditional methods, reliant on growth-associated induction, is non-trivial. Similarly, protein extraction methods are predominantly restricted to chemical methods, and mechanical methods reliant on expensive specialized equipment more tuned for large-scale applications. In this article, we outline detailed protocols for the use of an engineered autolysis/autohydrolysis E. coli strain, in two-stage fermentations in shake-flasks. This two-stage fermentation protocol does not require optimization of expression conditions and results in high protein titers. Cell lysis in an engineered strain is tightly controlled and only triggered post-culture by addition of a 0.1% detergent solution. Upon cell lysis, a nuclease digests contaminating host oligonucleotides, which facilitates sample handling. This method has been validated for use at different scales, from microtiter plates to instrumented bioreactors.
Graphic abstract:
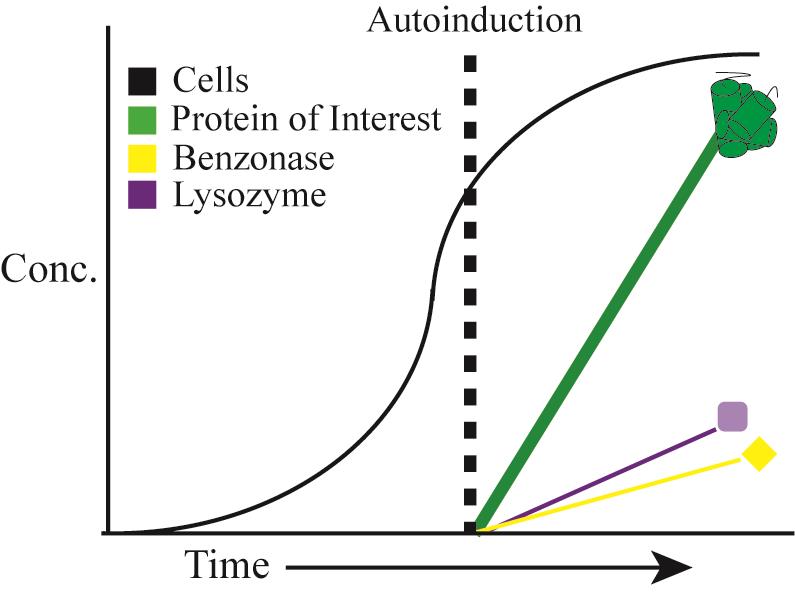
Two-stage protein expression, cell autolysis and DNA/RNA autohydrolysis. Reprinted with permission from Menacho-Melgar et al. (2020a). Copyright 2020 John Wiley and Sons.
Background
Despite heterologous protein expression being widely used in biotechnological research, optimization of protein expression protocols can be a very lengthy and costly process (Bill, 2014; Rosano and Ceccarelli, 2014). Once determined, optimal expression conditions are not readily translated to other fermentation scales or proteins of interest, thereby limiting method transfer to industrial scales, as well as increasing process development timelines (Mühlmann et al. 2017). After protein expression, many applications often require protein purification that calls for using expensive specialized equipment. Specifically, cell lysis for protein extraction is still mostly performed using mechanical methods, such as sonication or homogenization, which may not yield consistent results and limit high-throughput applications (Foster, 1992; Cai et al., 2008). Alternative methods, such as chemical methods orengineering “autolysis” microbial strains that lyse after cell harvesting, exist but are poorly controlled and do not account for oligonucleotides removal to reduce sample viscosity, which can complicate sample handling (Chien and Lee, 2006; Cai et al., 2008).
To address these, we recently described a two-stage protein expression process (Menacho-Melgar et al., 2020b). In this method, we use engineered strains that do not produce organic acid byproducts (e.g., acetate), known to negatively affect cell growth and protein expression (Jian et al., 2010), in combination with plasmids, where the expression of the protein of interest is under the tight control of a low phosphate inducible promoter (refer to Figure 1 for an example annotated plasmid sequence) (Moreb et al., 2020). In two-stage fermentations, protein expression is triggered upon entry into the stationary phase, when phosphate concentration in the media becomes depleted, which (i) stops cell growth (stage 1) and (ii) induces protein expression (stage 2) (Figure 2A). Two-stage fermentations decouple cell growth from protein expression, which results in high protein titers and enables the use of already optimized standard protocols across different scales and proteins of interest (Burg et al., 2016; Decker et al., 2020). In other words, two-stage protein expression does not require optimizing induction conditions, as opposed to methods that induce protein expression during growth. Additionally, this approach has enabled higher protein titers than growth-associated protein expression.
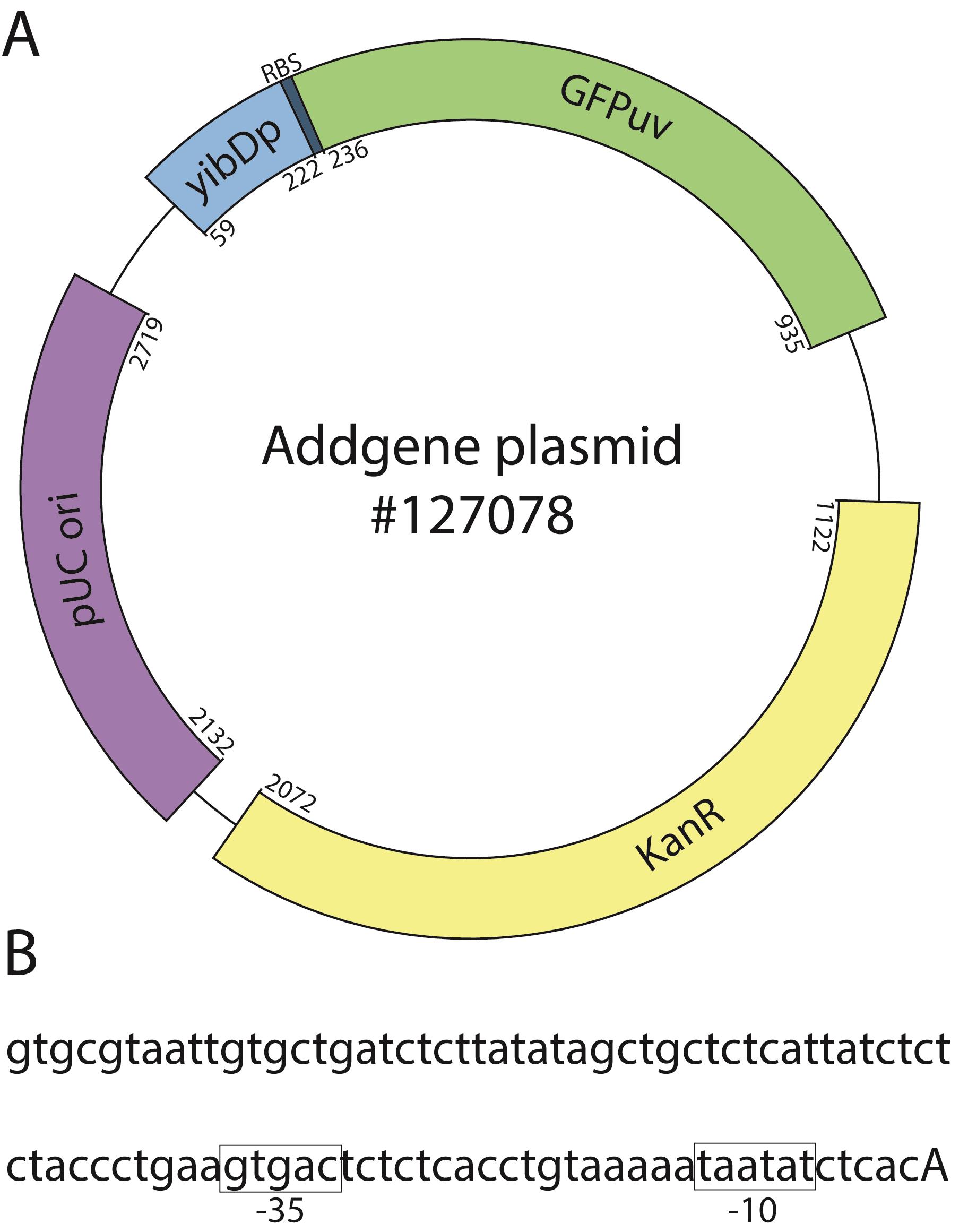
Figure 1. Example two-stage expression plasmid. A. Expression of the protein of interest (GFPuv, green) is driven by a low-phosphate inducible promoter (yibDp, blue). The plasmid has the high copy pUC origin (pBR322 derivative, purple) and a kanamycin resistance marker (yellow). B. Partial yibD promoter sequence showing the -10 and -35 boxes (highlighted in boxes) and the start of transcription (capitalized). Start and end bases for each feature are annotated. For a complete plasmid sequence, see https://www.addgene.org/browse/sequence/271197/ under the ‘Sequence’ tab.
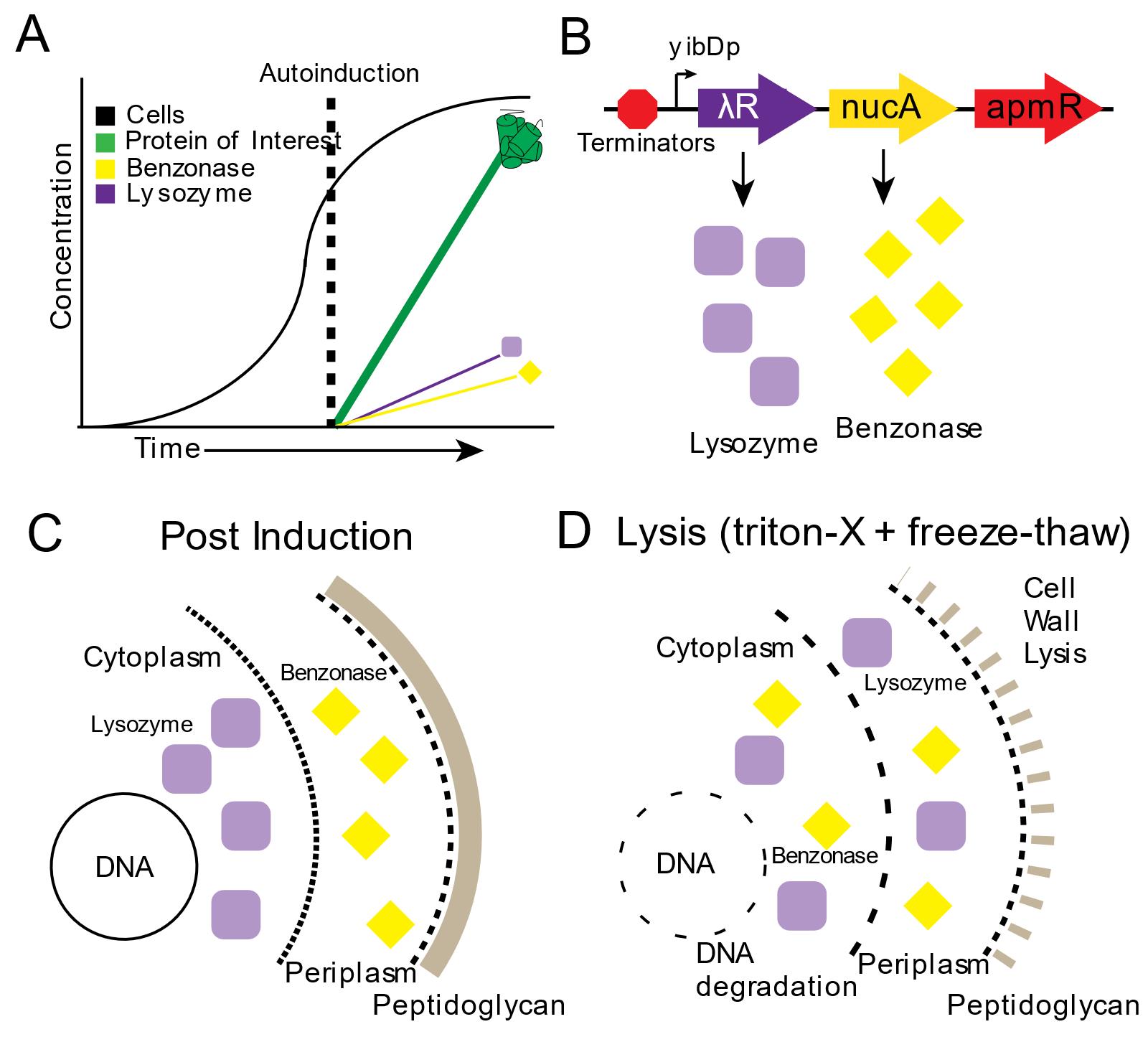
Figure 2. Autolysis and Autohydrolysis in two-stage protein expression. A. In stage 1 “growth”, cells are grown in AB-2 media. In stage 2, “production” gets triggered upon phosphate depletion in the media, when cells enter the stationary phase, activating expression of the protein of interest. B. Expression of the autolysis/autohydrolysis machinery, that is chromosomally integrated and consists of benzonase and lysozyme, also occurs at this stage. C. After induction, lysozyme remains inside the cytoplasm and benzonase is located in the periplasmic space. D. Only upon addition of detergent (Triton-X) and a freeze-thaw cycle, the cells lyse and the endonuclease is able to digest the host’s DNA. Reprinted with permission from Menacho-Melgar et al. (2020a). Copyright 2020 John Wiley and Sons.
Additionally, we built upon two-stage protein expression and engineered an autolysis/autohydrolysis strain, which expresses a lysozyme (from the lambda R gene) in the cytoplasm and a nuclease (from S. marcescens nucA gene, commercially known as BenzonaseTM) in the periplasm, during the protein production stage (Figure 2A-2C) (Menacho-Melgar et al., 2020a). These chromosomally integrated enzymes do not negatively affect the titer of the protein of interest (Menacho-Melgar et al., 2020a). Since this system is tightly controlled, no cell lysis will occur during the growth phase or after harvesting, as opposed to other BL21-based autolysis strains (Menacho-Melgar et al., 2020a). Only upon resuspension in a lysis buffer containing 0.1% detergent and a freeze-thaw cycle, over 90% of the protein is released (compared to a 20-minute sonication protocol, where further sonication does not lead to more protein release) and all of the oligonucleotides in the cell lysate are removed (Figure 2D) (Menacho-Melgar et al., 2020a). Importantly, this method enables the simultaneous lysis of several samples, which can be essential in high-throughput applications (Menacho-Melgar et al., 2020a). Although this method has been validated across different fermentation scales (Menacho-Melgar et al., 2020a), fermentations in microtiter plates and bioreactors require specialized equipment and accessories not readily accessible, whereas fermentations in shake-flasks are more widely and routinely used. Thus, in this protocol, we have described a detailed protocol for performing two-stage fermentations only in shake flasks, as well as protein extraction using our autolysis/autohydrolysis strain and a plasmid expressing GFPuv (Addgene, #127078), as an example (Menacho-Melgar et al., 2020a).
Materials and Reagents
10, 200, and 1,000 μL pipette tips (Genesee Scientific, catalog numbers: 23-121RLC, 23-150RLC and 23-165RLC)
1.7 mL microcentrifuge tubes (Genesee Scientific, catalog number: 24-281)
15 mL culture tubes (Genesee Scientific, catalog number: 21-130)
50 mL conical tubes (Genesee Scientific, catalog number: 28-108)
2 mL cryovials (VWR, catalog number: 10018-754)
1 L media bottles (VWR, catalog number: 10754-820)
10 mL and 25 mL serological pipets (Genesee Scientific, catalog numbers: 12-104 and 12-106)
250 vented baffled shake flask (VWR, catalog number: 89095-270)
Tryptone (Biobasic, catalog number: TG217(G211))
Yeast extract (Biobasic, catalog number: G0961)
Sodium chloride (Biobasic, catalog number: DB0483)
Ammonium sulfate anhydrous (VWR, catalog number: M105)
Glucose (VWR, catalog number: 89405-376)
Bis-Tris (GoldBio, catalog number: B-020-500)
Casamino acids (Biobasic, catalog number: CB3060)
12 M Hydrochloric acid (Biobasic, catalog number: HC6025)
Tris base (GoldBio, catalog number: T-400-500)
Triton X-100 (Sigma Aldrich, catalog number: 93443)
Protease inhibitors tablets EDTA-free (A32965)
Strain DLF_R004 (F-, λ-, Δ(araD-araB)567, lacZ4787(del)::rrnB-3), rph-1, Δ(rhaD-rhaB)568, hsdR51, ΔackA-pta, ΔpoxB, ΔpflB, ΔldhA, ΔadhE, ΔicIR, ΔarcA, ΔompT::yibDp- λR-nucA-apmR) transformed with a plasmid expressing your protein of interest (e.g., GFPuv), under a low phosphate inducible promoter (e.g., yibD) (e.g., Addgene plasmid #127078)
Low salt LB (see Recipes)
AB-2 (see Recipes)
Lysis buffer (see Recipes)
Equipment
Pipette controller (Eppendorf, model: 4430 000 018)
Incubator shaker, orbit set to 50 mm (Kuhner, model: ISF4-X)
Benchtop centrifuge (Thermo Scientific, model: Legend XTR)
-80°C freezer (Thermo Scientific, model: TSX700)
Microcentrifuge (Eppendorf, model: 5415R)
Heat block (Bioer, model: CHB-201)
Procedure
Shake-flask two-stage fermentations
Inoculate a 15 mL culture tube containing 5 mL of sterile low salt LB supplemented with appropriate antibiotics, using a frozen stock of DLF_R004 expressing your protein of interest (e.g., if using Addgene #127078 plasmid expressing GFPuv, a concentration of 35 µg/mL of kanamycin sulfate is needed).
Incubate the 5 mL cell culture at 37°C overnight at 150 rpm (shaker orbit: 50 mm).
Fill each 250 mL shake flask with 20 mL of sterile AB-2, supplemented with appropriate antibiotics.
Inoculate the AB-2 media with the overnight LB culture at 1% v/v (using 200 μL of the LB overnight for each 250 mL shake flask).
Incubate the AB-2 culture(s) for 24 h at 37°C with shaking at 150 rpm (shaker orbit: 50 mm)
After 24 h, since protein expression has finished (Menacho-Melgar et al., 2020b), measure and record the OD. While we have observed protein expression above OD 5, typical OD values range from 10 to 20. If an OD of 10 has not been reached, please refer to ‘Adaptation to alternative cell culture set-ups” under ‘Notes’ below.
Harvest the cells by centrifugation at 4,000 × g for 20 min.
Protein extraction using autolysis/autohydrolysis (refer to Figure 3)
Resuspend the cells to one-tenth of the original cell culture volume in 1× Autolysis buffer (2 mL total volume per 20 mL shake flask culture) supplemented with protease inhibitors, according to the manufacturer’s recommendations. For ease of handling, transfer the cell resuspensions from shake-flasks to microcentrifuge tubes.
Freeze the cells for 1 h in a -20°C or -80°C freezer. Ensure the cells become frozen, by tilting the tube and checking the cells do not flow. This is a good stopping point for this protocol, so one can use longer freezing times (e.g., overnight) without affecting the lysis.
Thaw the cells at 37°C in a heat block and incubate them for 1 h. Incubation at a lower temperature (e.g., room temperature) will lead to incomplete cell lysis.
Centrifuge the cells at 15,000 × g in a microcentrifuge.
Collect the supernatant containing the DNA/RNA-free protein extract.
To quantify protein titer, we recommend using a Bradford assay (to measure total protein concentration) coupled with an SDS-PAGE (to measure expression levels).
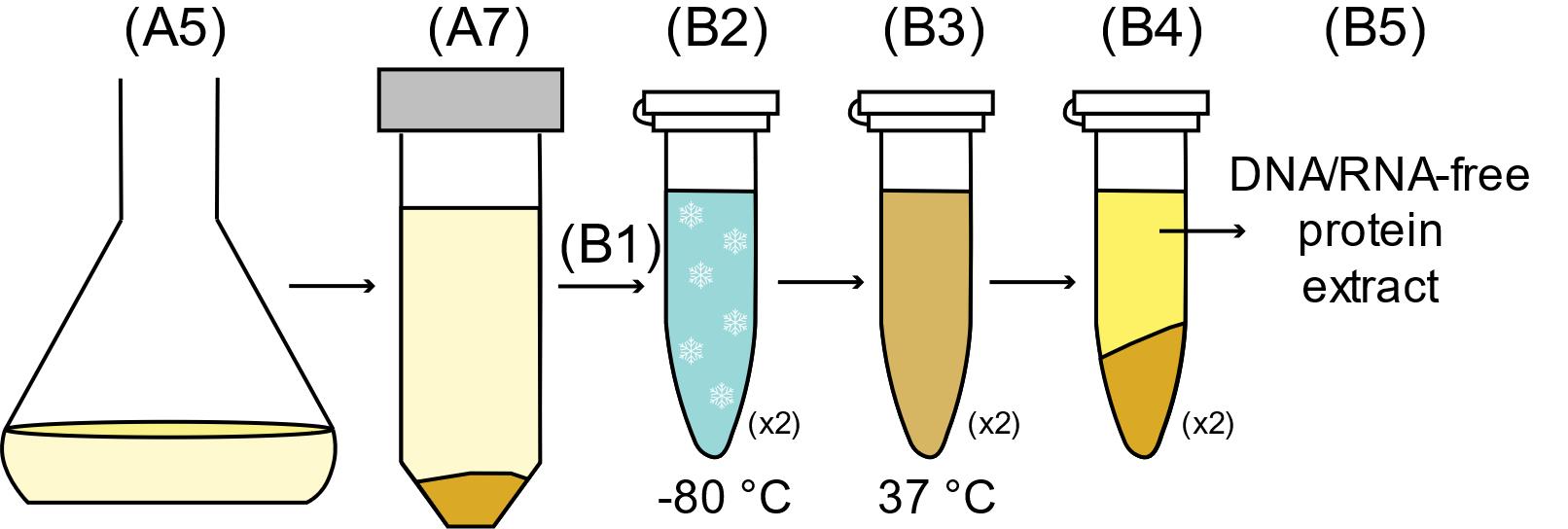
Figure 3. Autolysis/Autohydrolysis protocol outline. After growing the cells in 20 mL of AB-2 in a shake-flask for 24 hours (Step A5), the cells are harvested by centrifugation (Step A7), resuspended in 1x Lysis buffer (Step B1, two tubes for a single shake-flask), and frozen for at least 1 h (Step B2). The cells are thawed and incubated at 37°C for 1 h (Step B3). The lysate is then centrifuged (Step B4) and the protein extract can be collected (Step B5).
Notes
Adaptation to alternative cell culture set-ups.
Differences in equipment or materials may affect the performance of this protocol. If said variations exist, and the cell incubation conditions indicated in this protocol do not result in high protein expression (>5% expression level, visible in an SDS-PAGE gel) and complete lysis, we recommend doing an optimization cycle, where fill volumes and the shaking rpm are varied to determine the optimal conditions for your setup. For GFPuv, the protein titer should be 1.9 g/L or ~40 mg per shake flask.
Recipes
Low salt LB (Lennox formulation)
10 g of tryptone
5 g of sodium chloride
5 g of yeast extract
Adjust to 1,000 mL of deionized water and autoclave.
AB-2
Solution 1:
500 g of glucose
Fill to 1,000 mL with deionized water and autoclave.
Solution 2:
41.8 g of Bis-Tris
5.4 g of ammonium sulfate
6.2 g of yeast extract
3.5 g of casamino acids
3 mL of HCl
Fill to 910 mL with deionized water and autoclave
For 1 L of AB-2, add 90 mL of sterilized Solution 1 to 910 mL of sterilized Solution 2.
Lysis buffer
20 mM Tris HCl, pH 8.0
0.1% v/v Triton X-100
Add 1 tablet of protein inhibitors per 50 mL of lysis buffer.
Acknowledgments
We would like to acknowledge the following support: DARPA# HR0011-14-C-0075, ONR YIP #N00014-16-1-2558, and DOE EERE grant #EE0007563, the North Carolina Biotechnology Center 2018-BIG-6503 and NIH R61 AI140485-01.
Competing interests
MD Lynch has a financial interest in DMC Biotechnologies, Inc. MD Lynch and R Menacho-Melgar have financial interests in Roke Biotechnologies, LLC.
References
- Bill, R. M. (2014). Playing catch-up with Escherichia coli: using yeast to increase success rates in recombinant protein production experiments. Front Microbiol 5: 85.
- Burg, J. M., Cooper, C. B., Ye, Z., Reed, B. R. and Lynch, M. (2016). Large-scale bioprocess competitiveness: the potential of dynamic metabolic control in two-stage fermentations. Curr Opin Chem Engi 14: 121-136.
- Chien, L. J. and Lee, C. K. (2006). Synergistic effect of co-expressing D-amino acid oxidase with T7 lysozyme on self-disruption of Escherichia coli cell. Biochem Eng J 28(1): 17-22.
- Cai, Z., Xu, W., Xue, R. and Lin, Z. (2008). Facile, reagentless and in situ release of Escherichia coli intracellular enzymes by heat-inducible autolytic vector for high-throughput screening. Protein Eng Des Sel 21(11): 681-687.
- Decker, J. S., Menacho-Melgar, R. and Lynch, M. D. (2020). Low-Cost, Large-Scale Production of the Anti-viral Lectin Griffithsin. Front Bioeng Biotechnol 8: 1020.
- Foster, D. (1992). Cell Disruption: Breaking Up is Hard to do. Bio/Technology 10(12): 1539.
- Jian, J., Zhang, S. Q., Shi, Z. Y., Wang, W., Chen, G. Q. and Wu, Q. (2010). Production of polyhydroxyalkanoates by Escherichia coli mutants with defected mixed acid fermentation pathways. Appl Microbiol Biotechnol 87(6): 2247-2256.
- Mühlmann, M., Forsten, E., Noack, S. and Büchs, J. (2017). Optimizing recombinant protein expression via automated induction profiling in microtiter plates at different temperatures. Microb Cell Fact 16(1): 220.
- Menacho-Melgar, R., Moreb, E. A., Efromson, J. P., Yang, T., Hennigan, J. N., Wang, R. and Lynch, M. D. (2020a). Improved two-stage protein expression and purification via autoinduction of both autolysis and auto DNA/RNA hydrolysis conferred by phage lysozyme and DNA/RNA endonuclease. Biotechnol Bioeng 117(9): 2852-2860.
- Menacho-Melgar, R., Ye, Z., Moreb, E. A., Yang, T., Efromson, J. P., Decker, J. S., Wang, R. and Lynch, M. D. (2020b). Scalable, two-stage, autoinduction of recombinant protein expression in E. coli utilizing phosphate depletion. Biotechnol Bioeng 117(9): 2715-2727.
- Moreb, E. A., Ye, Z., Efromson, J. P., Hennigan, J. N., Menacho-Melgar, R. and Lynch, M. D. (2020). Media Robustness and Scalability of Phosphate Regulated Promoters Useful for Two-Stage Autoinduction in E. coli. ACS Synth Biol 9(6): 1483-1486.
- Rosano, G. L. and Ceccarelli, E. A. (2014). Recombinant protein expression in Escherichia coli: advances and challenges. Front Microbiol 5: 172.
Article Information
Copyright
© 2022 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Menacho-Melgar, R. and Lynch, M. D. (2022). Simple Scalable Protein Expression and Extraction Using Two-stage Autoinducible Cell Autolysis and DNA/RNA Autohydrolysis in Escherichia coli . Bio-protocol 12(2): e4297. DOI: 10.21769/BioProtoc.4297.
Category
Microbiology > Heterologous expression system > Escherichia coli
Molecular Biology > Protein > Expression
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.