- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

A Combinatorial in-silico, in-vitro and in-vivo Approach to Quantitatively Study Peptide Induced MHC Stability
Published: Vol 11, Iss 24, Dec 20, 2021 DOI: 10.21769/BioProtoc.4255 Views: 4471
Reviewed by: Luis Alberto Sánchez VargasMarie BoutetRAMESH KUDIRA
Original research article
The authors used this protocol in:

Apr 2021

Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Abstract
Here, we describe a combinatorial approach in reverse vaccinology to identify immunogenic class I major histocompatibility complex (MHC) displayed epitopes derived from a morbillivirus named pestes des petits ruminants (PPRV). The protocol describes an in silico prediction of immunogenic epitopes using an IEDB tool. The predicted peptides were further analysed by molecular docking with mouse class I MHC (H-2Kb), to assess their binding affinity, and their immunogenicity was validated, using acellular and cellular assays. Finally, an enumeration of the expanded PPRV-specific CD8+ T cells in infected or immunized mice against the immunogenic peptides was performed ex vivo. Synthetic peptide derivatives from different structural and non-structural proteins of PPRV were used to measure the extent of stabilized H2-Kb, using an ELISA based acellular assay and TAP deficient RMA/s cells. Fluorescently labelled H2-Kb-tetramers were generated by displacing a UV photocleavable conditional ligand with the PPRV-peptides. The resulting reagents were used to identify and enumerate virus-specific CD8+ T cells in immunized or PPRV-infected mice. The combinatorial approach described here could be used to identify immunogenic epitopes of any pathogen, autoantigens, as well as cancer antigens.
Graphic abstract:
Figure 1. General schematic to identify immunogenic peptides and their stabilization on MHC I molecule.
Background
Major histocompatibility complex (MHC) molecules loaded with peptides are displayed on the cell surface to engage T cells via their expressed TCRs. While all nucleated cells express class I MHC molecules, professional APCs such as dendritic cells, macrophages, and B cells additionally express high level of class II MHC molecules to activate T cells (Wieczorek et al., 2017). Formation of assembled class I and II MHC molecules requires binding of the proteolytically generated peptides with specific positioning of anchor residues. Endogenously synthesized proteins or those acquired exogenously by APCs are processed in distinct subcellular compartments to generate peptides that are presented by class I MHC molecules at the cell surface (Princiotta et al., 2003). Endogenous proteins are primarily processed by proteasomal machinery, and peptides thus generated are translocated from cytosol to endoplasmic reticulum (ER) mainly by the transporter associated with antigen processing (TAP) molecules (Vyas et al., 2008). Nascent class I MHC molecules in the ER associate with calreticulin, tapasin, and ERp57 to generate a peptide loading complex (PLC) that facilitates loading of peptides. Then, the assembled complexes traverse to the cell surface. Therefore, TAP deficiency limits the entry of peptides into ER, leading to a reduced surface expression of class I MHC molecules. The heterotrimeric complex formed by heavy chain (HC) of class I MHC molecule, peptide, and β2 microglobulin (β2m) provides the central anchorage signal to engage specific TCR expressing cytotoxic T lymphocytes (CTLs). Additionally, the co-stimulatory receptor-ligand pair (B7.1/7.2 and CD28) by the engaged APCs and T cells helps to efficiently activate CTLs, which are then enormously expanded in the presence of generated cytokine milieu (De Bruijn et al., 1991; La Gruta et al., 2018). Such cells scan infected or transformed cells for cytolysis. Therefore, identifying pathogen derived peptides constitutes a critical step in understanding T cell dynamics following infection or immunization. To validate the immunogenicity of the predicted peptides, fluorescently labelled H-2Kb/peptide complexes were generated and used to detect PPRV-peptides specific CD8+ T cells in immunized or the PPRV-infected mice.
In this protocol, we describe assays to ascertain peptide induced class I MHC stabilization (Wieczorek et al., 2017). A combinatorial approach involves in silico, in vitro and in vivo analysis to predict immunogenic PPRV-peptides that can be displayed by class I MHC molecules of mice (H-2Kb). Finally, a throughput approach was used to enumerate PPRV-specific CD8+ T cells in the immunized or infected mice. For identifying such peptides, we collected the complete amino acid sequences of PPRV proteins from online NCBI and Uniprot databases to predict class I MHC restricted epitopes using online tools such as IEDB, SYFPEITHI (Figure 1). High affinity epitopes were manually selected based on their hydrophobic residues at carboxy terminus, as the latter are known to favor peptide-MHC class I interaction (Pettersen et al., 2004; Chen et al., 2015). Furthermore, the selected high affinity peptides were subjected to a blind docking analysis with H-2Kb, whereby the top ranked peptides were selected (Vita et al., 2015 and 2019; Zhou et al., 2018). To study the presentation of such peptides in context to H-2Kb, we used TAP deficient RMA/S cells that inherently express low levels of surface class I MHC molecules. RMA/s cells can synthesize heavy chains of class I MHC molecules and β2m, but fail to efficiently assemble these molecules due to the limited availability of processed peptide in the ER (Esquivel et al., 1992; De Silva et al., 1999). This leads to compromised loading of class I MHC molecules with antigenic peptides. Empty class I MHC molecules can, nonetheless, be transported to the cell surface by vesicular transport from the ER (De Bruijn et al., 1991; Esquivel et al., 1992). Empty class I MHC molecules are poorly stabilized at the cell surface and hence lower expression levels are evident. Exogenously added antigenic peptides to RMA/S cells can be loaded to class I MHC molecules by predominantly two mechanisms. First, the exogenous peptides can bind directly to the empty class I MHC molecules at the cell surface (Schumacher et al., 1990). Second, the peptides are internalized by such cells, loaded onto the peptide binding grove of heavy chain (HC) of class I MHC molecule utilizing the vacuolar pathway to generate stable heterotrimeric complexes involving β2m, and subsequently the complexes are efficiently displayed on the cell surface (Figure 2) (Esquivel et al., 1992; De Silva et al., 1999). Fine details of such processes are still to be better defined. The binding ability of anchor residues of the peptide with MHC molecules dictates the stability of such complexes (Garstka et al., 2015). We showed that stable class I MHC molecules were expressed on the cell surface when immunogenic peptides derived from PPRV were added to RMA/s cells, and such molecules were detected by anti-H-2Kb antibody using flow cytometry (Sharma et al., 2021). We also generated stable H-2Kb monomers using a photocleavable conditional ligand, (FAPG[Anp]YPAL), modified derivative from the nucleoprotein of Sendai E virus, which was described earlier byRodenko et al. (2006). Incubation of predicted PPRV-peptides in high concentration with such monomers, followed by exposure to a higher wavelength UV light (UV365), cleaved the conditional ligand and replaced it with the added immunogenic peptides (Rodenko et al., 2006). The exchange by different peptides was performed in separate identifiable wells of a microtiter plate, to unambiguously assess their MHC stabilization.
The described protocol can be employed to identify immunogenic epitopes of diverse antigens of a pathogen, cancers, as well as autoantigens; but the availability of cells deficient in antigen-processing machinery for the concerned species, as well as the generation of recombinant proteins for HC of class I MHC, and β2m would be critical. Furthermore, the final validation has to be performed by enumerating antigen-specific CD8+ T cells in the host. Therefore, a combinatorial approach could yield validated information on the immunogenicity of epitopes. Another limitation of such protocols might be the inherent polymorphism in MHC molecules occuring in an outbred population. Nonetheless, essential features of T cell differentiation are better captured if the information on immunogenic epitopes of an antigen is available. Further characterization of differentiating antigen-specific CD8+ T cells adds value to such analyses.
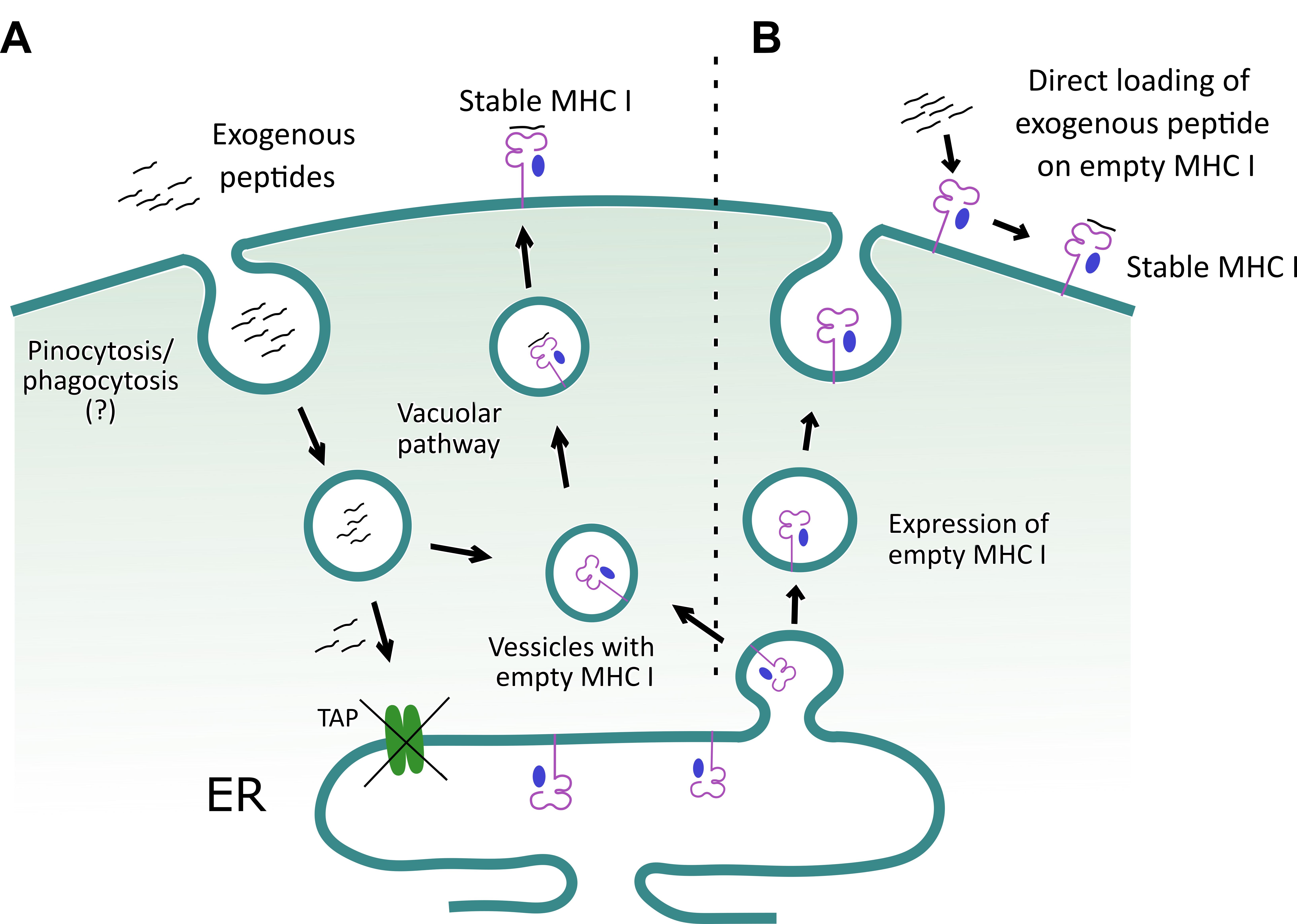
Figure 2. A schematic to depict how exogenous peptides help stabilize class I MHC molecule on the surface of TAP deficient RMA/S cells. A. Exogenous peptides are internalized and loaded onto class I MHC molecules in a yet to be clearly defined subcellular compartment. Subsequently, the loaded class I MHC complexes are exported to the cell surface. The surface stabilized class I MHC molecules are then detected by flow cytometry, using an anti-class I MHC (H-2Kb) antibody. B. Alternatively, RMA/s cells can display empty class I MHC molecule on their surface, and their loading with the exogenously added peptides helps to stabilize expression.
Materials and Reagents
1,000 µl tips (Tarsons, catalog number: 521020)
Racked filter tips, 1,000 µl (Tarsons, catalog number: 528104)
200 µl tips (Tarsons, catalog number: 521014)
Racked filter tips, 200 µl (Tarsons, catalog number: 528106)
10 µl tips (Tarsons, catalog number: 521000)
Racked filter tips, 10 µl (Tarsons, catalog number: 528100)
0.22 µm filtration units (Corning, catalog number: 431118)
5 L flasks (Borosil)
50 ml Centrifuge tubes (Tarsons, catalog number: 500041)
1.5 ml microcentrifuge tubes (Tarsons, catalog number: 500010)
Flat-bottom 96 well cell culture plate (Corning, catalog number: CLS3599-100EA)
Amicon concentrators (Merck, catalog number: UFC903024)
RMA/s cells (NCCS, Pune)
6-8 weeks old male and female C57BL/6 mice (Jackson Laboratories, catalog number: 000664)
H-2Kb and β2m plasmid constructs (Xu et al., 2016) (A kind gift from Hidde L. Ploegh Lab, Boston Children’s Hospital, Harvard University)
Peptides (GL Biochem, Shanghai)
Dimethyl sulfoxide, DMSO (Serva, catalog number: 20385.01)
D-(+)-Glucose anhydrous, Hi-ARTM/ACS (Himedia, catalog number: GRM077-500G)
Ampicillin sodium salt (Himedia, catalog number: MB104-5G)
Isopropyl β-d-1-thiogalactopyranoside, IPTG (BR Biochem Life sciences, catalog number: BC0168-100G)
Lysozyme (Himedia, catalog number: MB098-5G)
DNase I (Sigma-Aldrich, catalog number: 11284932001)
Magnesium Chloride, MgCl2 (Merck, catalog number: 105833)
Ethylene diamine tetraacetic acid, EDTA (Merck, catalog number: 324503)
Sodium-Azide (Merck, catalog number: 8223350100)
Tris-Hydrochloride, Tris-HCl (Himedia, catalog number: GRM1218-5KG)
Sucrose (Duchefa Biochemie, catalog number: S0809.1000)
Diothiothriotol, DTT (Himedia, catalog number: RM525-5G)
Triton X-100 (Serva, catalog number: 37240.01)
Tween 20 (Himedia, catalog number: MB067-100ML)
Sodium Chloride, NaCl (Himedia, catalog number: MB023-500G)
Sodium Hydroxide, NaOH (Himedia, catalog number: MB095-500G)
Di-sodium hydrogen phosphate, Na2HPO4 (Himedia, catalog number: GRM3960-500G)
Potassium dihydrogen phosphate, KH2PO4 (Himedia, catalog number: GRM1188: 500G)
Potassium Chloride, KCl (Merck, catalog number: 1049330500)
Urea (Himedia, catalog number: MB032-5KG)
L-Arginine (Sigma-Aldrich, catalog number: A5006)
Oxidised Glutathione (Himedia, catalog number: RM550-500MG)
Reduced Glutathione (Himedia, catalog number: RM234-5G)
Guanidine-hydrochloride (Serva, catalog number: 39558.02)
Sodium acetate, C2H3NaO2 (Himedia, catalog number: MB048-500G)
Ammonium Chloride, NH4Cl (Merck, catalog number: 1011450500)
Sodium bicarbonate, NaHCO3 (Serva, catalog number: 30180.02)
Polymethylsulphonyl fluoride, PMSF (Serva, catalog number: 32395.04)
4-Nitrophenyl phosphate disodium salt hexahydrate (Sigma-Aldrich, catalog number: 71768-25G)
Propidium iodide, PI (BD biosciences, catalog number: 556463)
Luria Bertani broth, LB broth, Miller (Himedia, catalog number: M1245-1KG)
LB media broth components include:
Tryptone
Yeast extract
Sodium Chloride
BirA biotin-protein ligase standard reaction kit (Avidity)
Kit components include:
d-Biotin (500 µM)
Biotin ligase BirA enzyme (1 mg/ml)
Solution A (bicine buffer: 0.5 M bicine, pH 8.3)
Solution B (ATP biotin: 100 mM ATP, 100 mM MgOAc, 200 µM biotin)
Purified Streptavidin (ThermoFisher, catalog number: SNN1001)
APC conjugated Streptavidin (Biolegend, catalog number: 405207)
RPMI (Gibco, catalog number: 31800-022)
Fetal bovine serum, FBS (Gibco, catalog number: 26140-079)
Penicillin-Streptomycin (Gibco, catalog number: 10378-016)
Anti-mouse FITC-H-2Kb antibody (BD Pharmingen, Clone: AF6-88.5, catalog number: 562002)
Anti-Mouse IgG (whole molecule)-Alkaline phosphatase antibody produced in goat (Sigma-Aldrich, catalog number: A9316-1ML)
Anti-mouse CD8 antibody (clone 53-6.7, BD Bioscience)
Freund’s Adjuvant, complete (Sigma-Aldrich, catalog number: F5881-10ml)
Freund’s Adjuvant, incomplete (Sigma-Aldrich, catalog number: F5506-10ml)
PPRV vaccine strain Sungri/96
Stock solutions (see Recipes)
LB media broth (see Recipes)
Phosphate buffered saline, 1× PBS (see Recipes)
PBS-T (see Recipes)
FACS Buffer (see Recipes)
Resuspension buffer (see Recipes)
Lysis buffer (see Recipes)
Washing buffer with Triton (see Recipes)
Washing buffer without Triton (see Recipes)
Urea buffer (see Recipes)
Refolding buffer (see Recipes)
Guanidine solution (see Recipes)
Gel filtration buffer (see Recipes)
Red blood cell (RBC) Lysis Buffer (see Recipes)
Equipment
Serological and micropipettes (Eppendorf Research Plus)
Spectrophotometer (GeneQant, model: 80-2130-00)
Autoclave (ALP, model: CL-40MDP)
Incubator with shaker (TBOY 5000I INCUB SHKER, catalog number: 980473)
Sonicator (Sonics & Materials INC., model: VCX750)
Centrifuges (Eppendorf, models: AG 5810 R and 5401)
Gel filtration apparatus (AKTA) with cold chamber (HiPrep 16/60, Sephacryl S200 HR)
UV-chamber (Genetix, model: GX-2082-6111)
BD Accuri C6 Flow cytometer (BD Biosciences)
SDS gel apparatus (Bio Rad Mini-PROTEAN Tetra System)
Eon ELISA Plate Reader (BioTek Instruments, Inc.)
LSE Vortex Mixer (Corning, catalog number: 6776)
Laboratory Stirrer/Hot Plate (Corning, catalog number: 6798-420D)
Magnetic bars (Tarsons, catalog number: 4117)
Software
UCSF Chimera version 1.13 (https://www.cgl.ucsf.edu/chimera) (Pettersen et al., 2004; Chen et al., 2015)
HPEPDOCK online docking tool (http://huanglab.phys.hust.edu.cn/hpepdock/) (Zhou et al., 2018)
NCBI, PDB, Uniprot protein Databases (https://www.ncbi.nlm.nih.gov/; https://www.rcsb.org/; https://www.uniprot.org/)
Immune Epitope Database and Analysis Resource (IDEB) and IDEB-AR (Zhang, 2013)
FlowJo vX (Becton Dickinson and company)
GraphPad Prism v8.4.3 (San Diego, CA)
Microsoft office 2016
Procedure
Prediction of immunogenic epitopes of PPRV using bioinformatic tools (Figure 1)
Obtain amino acid sequence of different PPRV proteins from various databases, such as National Centre for Biotechnology Information (NCBI), Protein Data bank (PDB), UniProt, Structural Classification of Proteins (SCOP), and CATH: Protein Structure Classification database. It can be done by searching the protein by its name in the respective databases and downloading the desired sequence in Fasta Format. For example, the sequence of H protein of PPRV is obtained by selecting protein database of NCBI and the input query is ‘PPRV H protein’. Then select the hit with 609 amino acids, a putative complete amino acid sequence for the H protein, and copy the sequence in Fasta format.
Use open access Immune Epitope Database (IEDB) or IDEB online database to insert the sequences of the viral proteins as the query (Vita et al., 2015 and 2019).
Customize the peptide lengths for mouse class I MHC molecule (H-2Kb), and predict immunogenic epitopes.
Bin the predicted epitopes according to the binding affinity to H-2Kb, and select the epitopes with percentile ranks of <2 and IC50 values lower than 50 nM, which are considered high affinity binders.
Validate the high affinity binders using IEDB-AR (a companion site to IEDB that provides multiple options for analyzing the predicted T cell epitopes), and further scrutinize them on the basis of general characteristics of naturally occurring immunogenic peptides such as the 9mer, FAPGNYPAL of SEV complexed with H-2Kb.
Select the top ranking 9mers (as these are the most naturally occurring high affinity class I MHC restricted accessible epitopes) and scrutinize them for the presence of hydrophobic carboxy terminal residues.
Rank the selected peptides based on the affinity scores, and obtain commercially synthesized peptides with free N and C termini, and high purity (95-98%). Use them further for experimental analysis.
Molecular docking of the peptides with class I MHC molecules for determining binding affinities (Figure 3)
Perform molecular docking of high affinity epitopes with H-2Kb (in PDB file), using their individual sequence as the input for homology modelling by a HPEPDOCK online server tool (e.g., file 1S7Q for modelling of PPRV epitopes with H-2Kb Protein) (Zhou et al., 2018).
The server predicts and generates an ensemble of docked structures. Analyze the energy parameters for the docked structures of H-2Kb and peptide. Set a cut-off value to select peptides based on those of the known epitopes of similar length with the H2-Kb (e.g., SEV nucleoprotein derived peptide, FAPGNYPAL for 9mers and chicken ovalbumin peptide, SIINFEKL for 8mers). Select the docking structures with energy parameters lower than or equal to the standard epitopes.
Further scrutinize the selected docking structures based on the following parameters:
N terminus of SEV-9 peptide is deeply embedded into H2-Kb, while C-terminus is held by salt-bridge near the surface of H-2Kb.
Residues 4 or 5 of SEV-9 peptide are bulging out of the groove and not interacting with H-2Kb via Van der Waal’s forces.
Polar interactions of peptide termini with H-2Kb stabilize the SEV-9 peptide.
Residues 5 or 6 serve as the most probable primary anchor residues, while amino acids at positions 8 or 9 might act as the secondary anchors in the SEV-9 Peptide.
All the parameters are documented in a tabular form to select the most stable docking structures.
Repeat Step B3 for all the docked structures.
Select the most stable docked structures from each of the high affinity epitopes, and compare their docking scores, i.e., docking energy and other parameters described in Step B3c. Plot a line curve or bar diagram to rank the epitopes according to immunogenicity and stabilizing H-2Kb.
Visualize the selected docked structures using UCSF chimera tool and binding analysis tool. Assess for class I MHC stabilization and finding contacts between class I MHC molecule and the predicted epitope.
Identify anchors based on the parameters described in Step B3.
Image docked structures using the pre-set option available in the chimera console. Use the same coloring scheme for all the structures and ribbon views for a better visualization.
Generation of heterotrimeric tetramers of H-2Kb (Figures 4 and 5)
To generate soluble H-2Kb/peptide complexes, overexpress H-2Kb heavy chain and β2m as recombinant proteins in E. coli, and refold the monomers in the presence of a high concentration of the conditional ligand FAPG(Anp)YPAL. The heavy chain sequence is mutated to remove the cytosolic and transmembrane regions, and is expressed with biotin acceptor peptide sequences at the C-terminus of the H-2Kb heavy chain, for site-specific labelling (Garboczi et al., 1992; Sehrawat et al., 2012; Pichugin et al., 2018). These short sequences can be easily biotinylated using BirA in a site-specific manner at lysine (K) residues. The four biotin binding sites of avidin/streptavidin then generate tetramers of H-2Kb/peptide complexes. To use these complexes for flow cytometry, different fluorochrome conjugated commercially available avidin/streptavidin can be used. In brief the following procedure is used.
Expression and purification of recombinant proteins (H-2Kb and β2m) from inclusion bodies.
Recombinant proteins of H-2Kb and β2m were purified from inclusion bodies collected from induced cultures of E. coli.
Prepare 6 Liter (L) of LB medium in 2 L flasks each with 1 L of medium. Autoclave these flasks and after cooling down at room temperature (RT), add 10 ml of sterile 40% glucose solution and 10 g of ampicillin to each flask.
Take 20 µl of bacterial stock and dilute it in 6 ml of LB medium; then, add 1 ml of the starter culture of bacterium to each flask.
Keep it in a shaker incubator at 37°C for approximately 8-10 h until the OD600 reaches 0.8. Intermittent sampling followed by measuring OD600 is suggested for capture growing bacteria in log phase.
Collect 1 ml of sample from each flask for the expression control.
When OD600 reaches 0.8, add 1 ml of freshly prepared IPTG solution (100 mg/ml in PBS, 1 mM) to individual flasks.
Incubate for 3 h at 37°C in a shaker incubator.
Cool down the cultures for 15-20 min on ice.
Harvest the bacterial cultures by pelleting 1 L cultures at 4°C by centrifuging at 11,000 × g for 10 min.
Resuspend the pellet in 13 ml of resuspension buffer, and transfer to a 50 ml centrifuge tube.
Homogenize the bacterial suspension in the resuspension buffer using a sonicator on ice (20% amplitude 15 s OFF/15 s ON pulse, total 30 cycles).
Add 100 µl of lysozyme (stock 50 mg/ml), 250 µl of DNase I (stock 2 mg/ml) and 50 µl of MgCl2 (stock 0.5 M).
Vortex thoroughly and add 12.5 ml of lysis buffer, followed by a brief vortexing.
Incubate at RT until viscosity decreases and then add 350 µl of Na·EDTA from 0.5 M stock.
Freeze and thaw the mixture by putting the tube in liquid nitrogen and then transferring it in a water bath at 37°C for 30 min. Add 200 µl MgCl2 and wait until viscosity decreases. Then add 350 µl Na EDTA and keep it on ice from now onwards.
Pellet the solution at 11,000 × g for 20 min at 4°C and discard the supernatant.
Resuspend the pellet in 10 ml of washing buffer with Triton X100 and homogenize again in a sonicator.
Pellet the solution at 11,000 × g for 20 min at 4°C and discard the supernatant.
Resuspend the pellet in 10 ml washing buffer without Triton X100 and homogenize using a sonicator.
Pellet down the mixture at 11,000 × g for 20 min at 4°C and discard the supernatant.
Collect the pellet and dissolve it in 5 ml of 8 M Urea; determine the protein concentration.
Make aliquots of 1 ml in separate vials and store at -80°C until future use.
Refolding of H-2Kb monomer with photo-cleavable peptide.
Prepare H2-Kb in 3 vials of 6.2 mg each (final concentration 3 µM) and β2m of 4.4 mg (final conc. 6 µM). All the preparation are diluted by guanidine solution up to 500 µl.
Cool down 200 ml of refolding buffer in a 500 ml glass flask on ice and add 2 ml of PMSF (stock 100 mM).
Dissolve 12 mg of photo-cleavable H-2Kb restricted peptide (stock 10 mg/ml in DMSO) in the refolding buffer.
Inject 1 vial of β2m dropwise to the refolding solution while this is being continuously vigorously stirred with the help of a magnetic bar on the rotating stirrer for rapid and efficient dilution.
Inject 1 vial of purified H-2Kb heavy chain drop by drop directly to the refolding solution.
Place the flask at 4°C for 8 h with slow stirring of the solution in a cold chamber.
Repeat Steps C2d to C2g three times.
After the third injection of β2m and H-2Kb, leave the flask with slow stirring for another 24 h.
After complete refolding of the solution, concentrate the protein up to 1 ml using a concentrator.
Load the concentrated protein in AKTA on a S200 column for gel filtration.
Collect different fractions of the refolded peak as revealed by 12% reducing SDS-PAGE. The fractions showing two distinct bands of ~35 kDa and 13 kDa are pooled and concentrated up to 1 ml (Figure 5).
In vitro biotinylation of H-2Kb/peptide complexes.
The in vitro biotinylation was done strictly as mentioned in the biotinylation kit (Avidity).
Add 100 µl of solution A and 100 µl of solution B to 700 µl of purified H-2Kb complexes.
Add 100 µl of d-biotin and 10 µl of BirA enzyme to the solution and incubate the mix overnight at RT.
Purify the biotinylated H-2Kb/peptide complex by running the sample on a S200 gel filtration column using AKTA.
Collect all the biotinylated fractions and concentrate the pooled volume to 1 ml or less.
Check the concentration of the biotinylated H-2Kb/peptide complex and aliquot in smaller volumes for storage.
For long term storage, add glycerol to a final concentration of 16%, make aliquots of 50 µl in 1.5 ml microcentrifuge tubes, and store them at -80°C for experimental uses and tetramerization.
Peptide-MHC binding assay using acellular assays (Figure 6)
Take an ELISA plate, coat it with 50 µl of streptavidin at a concentration of 2 µg/ml in PBS, and incubate at 4°C overnight.
Next day, wash the plates with 200 µl PBST to remove unbound streptavidin.
Block these wells with 200 µl of 3% BSA in PBS and incubate for 2 h at RT.
Remove the blocking buffer and wash with 200 µl PBST (three times).
Add 50 µl of biotinylated H-2Kb monomer (1 µg/ml) containing photo-cleavable peptide to the streptavidin coated wells and incubate for 1 h at RT.
Remove the unbound monomer and wash with 200 µl PBST (three times).
Add identified peptides to different wells, along with a positive control peptide (in this case we have used SIINFEKL). Label the wells appropriately.
Expose this plate in a chamber with high wavelength UV365 radiations for 30 min to achieve the displacement of UV-cleavable ligand with the respective peptides.
Wash these wells again with 200 µl PBST (three times).
Add 50 µl of mouse anti-β2 microglobulin antibody to all the wells for 1 h at RT.
Wash these wells again with 200 µl PBST (three times).
Add 50 µl of anti-mouse IgG antibody conjugated with alkaline phosphatase enzyme and incubate for 30 min at RT.
Wash these wells again with 200 µl PBST (three times).
Add 200 µl of p-nitrophenol phosphate (1 mg/ml) to the wells and wait for colour development due to the formation of a chromogenic product.
As soon as the color develops and is visible with eye, stop the reaction with 50 µl of NaOH 1M, and measure absorbance at 405 nm in a plate reader.
Take the readings and determine the best peptide that binds to H-2Kb.
Cell surface stabilization of MHC I using RMA/S Cell Lines (Figure 7)
RMA/S cells are defective in antigen processing due to a mutation in TAP molecules that enables peptide transport to the ER for its loading onto the newly synthesized class I MHC molecules. RMA/S cells display lower levels of empty class I MHC molecules that can be efficiently loaded with exogenous peptides (Schumacher et al., 1990; Esquivel et al., 1992). Peptides entering via pinocytosis or phagocytosis can also be efficiently loaded on empty class I MHC molecules by the vacuolar pathway. Therefore, the differential display of class I MHC molecules by RMA/s cells in the absence or presence of exogenous peptides can be utilized to measure the peptide induced stabilization of such molecules. To study MHC stabilization on RMA/S cells, the following procedure is used.
Revive RMA/s cells and culture them in RPMI media supplemented with 10% FBS in a humidified incubator with 5% CO2 at 37°C. Use RMA/S cells after three passages of sub-culturing to study the stabilization of class I MHC molecule.
Take 2 × 105 cells in 200 µl serum free RPMI media and starve these cells for 3 h at 37°C in a humidified incubator with 5% CO2.
Make different dilutions of peptides in serum free RPMI (Such as 6.25, 12.5, 25, 50 and 100 µg/ml).
After 3 h, wash these cells with 200 µl pre-warmed serum free RPMI, and add 200 µl of the prepared dilutions of peptides to the respective wells.
Incubate these cells for another 7-8 h in a humidified incubator at 37°C supplemented with 5% CO2.
Centrifuge the cells at 300 × g at RT and discard the supernatant.
Add 200 µl of FACS buffer and resuspend these cells.
Centrifuge the cells at 300 × g at RT and discard the supernatant.
Add 50 µl of FACS buffer and resuspend these cells.
Dilute H-2Kb antibody 1:200, add 50 µl to each well, and incubate for 20 min in the dark at RT. Keep three wells for staining control (H-2Kb only, PI only, and unstained).
Add 200 µl of FACS buffer and centrifuge at 300 × g for 5 min at RT. Repeat this step three times and resuspend the cells in 100 µl FACS buffer.
Prepare PI at a concentration of 2 µg/ml in FACS buffer and add 100 µl to all wells, except for the staining controls where PI is not required.
Instantly acquire the cells in a flow cytometer, and analyze the results using FlowJo software.
UV displacement of conditional ligand to generate fluorescent H-2Kb tetramers
Take 50 µl of concentrated biotinylated H-2Kb monomer (>250 µg/ml) refolded with the photo-cleavable peptide in a 96-well plate on ice, and dilute with gel filtration buffer to 100 µl.
Add 5 µl of specific peptides to each wells containing monomers with UV-cleavable ligand.
Expose these plates to UV radiation (365 nm) in a UV-chamber for 30 min on ice. The distance of the plate to the UV lamp should be 2-5 cm.
Centrifuge the plate at 3,000 × g for 5 min at 4°C, and collect 80 µl of the exchanged samples from the plate in microcentrifuge tubes.
Add 1 µl of streptavidin-APC conjugate in 19 µl of peptide exchanged monomer for tetramerization of the monomers; incubate for 2 h on ice.
Collect tetramers and use them to identify peptide specific CD8+ T cells, by co-staining with anti-CD8 antibody.
Detection of peptide specific CD8+ T cells in PPRV (Sunguri) infected mice (Figure 8)
Divide C57BL/6 WT mice in two groups with at least three animals per cage.
Infect mice of one cage intraperitoneally with a high dose of PPRV (5 × 106 pfu). The animals in the other cage will serve as control.
After 7 days, infect the same mice with a similar dose of PPRV virus to further expand the frequencies of PPRV specific cells.
Collect blood from each mouse from both groups and stain them to identify peptide specific CD8+ T cells in the blood.
Prepare a cocktail of PerCP Cy5.5 conjugated anti-mouse CD8 antibody (1:200 dilution) and peptide specific H-2Kb streptavidin-APC tetramer (1:100 dilution).
Transfer 5 µl of the cocktail to a fresh labelled microcentrifuge tube. Keep some tubes for staining control (e.g., CD8 control, H-2Kb-tetramer control, and unstained samples).
Add 25 µl blood from each mouse to the respective tubes and mix by gentle vortexing.
Incubate the tubes in the dark on ice for 30 min.
Add 220 µl of RBC lysis buffer to all the tubes, vortex gently, and wait for the RBCs to lyse.
After 5 min, add PBS and mix gently.
Instantly acquire the stained cells using a flow cytometer, and analyze for peptide specific CD8+ T cells.
Detection of peptide specific CD8+ T cells in the PPRV-peptide immunized mice
Place 600 µl of complete Freund’s adjuvant in a 1.5 ml microcentrifuge tube and add 6 µl of peptide (stock conc. 10 mg/ml).
Extensively vortex the tubes and collect the peptide emulsion in a 1 ml tuberculin syringe. Check the quality of emulsion by pouring a small drop of the emulsion in cold water. If the emulsion is good, as indicated by an intact drop, proceed with the injection to the mouse. If it is not good, keep the emulsion for another round of vigorous vortexing.
Inject 50 µl of the emulsion in each C57BL/6 WT mice subcutaneously, and wait for at least 15 days. Keep another three mice without any injection as control.
Prepare a fresh peptide emulsion in incomplete Freund’s adjuvant with the same volumes as in step 1, and subcutaneously inject in the previously immunised mice.
Collect blood samples from these mice at least 3 days after the second injection. The blood collected is used to detect peptide specific CD8+ T cells.
Prepare a cocktail of PerCP Cy5.5 conjugated anti-mouse CD8 antibody (1:200 dilution) and peptide specific H-2Kb streptavidin-APC tetramer (1:100 dilution).
Place 5 µl of the antibody cocktail in a fresh labelled microcentrifuge tube and keep some tubes for staining control (e.g., Anti-CD8 control, H-2Kb-tetramer control and unstained samples).
Add 25 µl blood from each mouse to the respective tubes, and mix by gentle vortexing.
Incubate the tubes in the dark on ice for 30 min.
Add 220 µl of RBC lysis buffer to all the tubes, vortex gently, and wait for the RBCs to lyse.
After 5 min, add PBS and mix gently.
Instantly acquire the stained cells using a flow cytometer, and analyze for peptide specific CD8+ T cells.
Data analysis
Prediction of viral peptides bioinformatically
After collecting the amino acid sequences in FASTA format from the NCBI database, immunogenic peptides were identified using the IEDB analysis resource. Around 80 peptides were predicted from this online server tool. These peptides were further scrutinized for high affinity binders. The IC50 values for high affinity binders were more than 500 nM. For predicting the binding affinities of these high percentile ranking peptides, molecular docking analysis was done. These peptides were exposed on the HPEPDOCK online server to obtain a peptide conformation when docked with H-2Kb protein. FAPGNYPAL was used as a positive reference to check the efficacy of the docking algorithm. We identified 12 immunogenic epitopes that satisfied all the parameters with high docking score (Figure 3A-3B). These predicted immunogenic peptides of different proteins of PPRV might have the potential to induce CD8 T cell response in mice. The best peptides are selected on the basis of their docking score to proceed with their stability on MHC I. An example of a predicted peptide that is analyzed bioinformatically using the HPEPDOCK online server tool is discussed briefly (Figure 3C-3D).
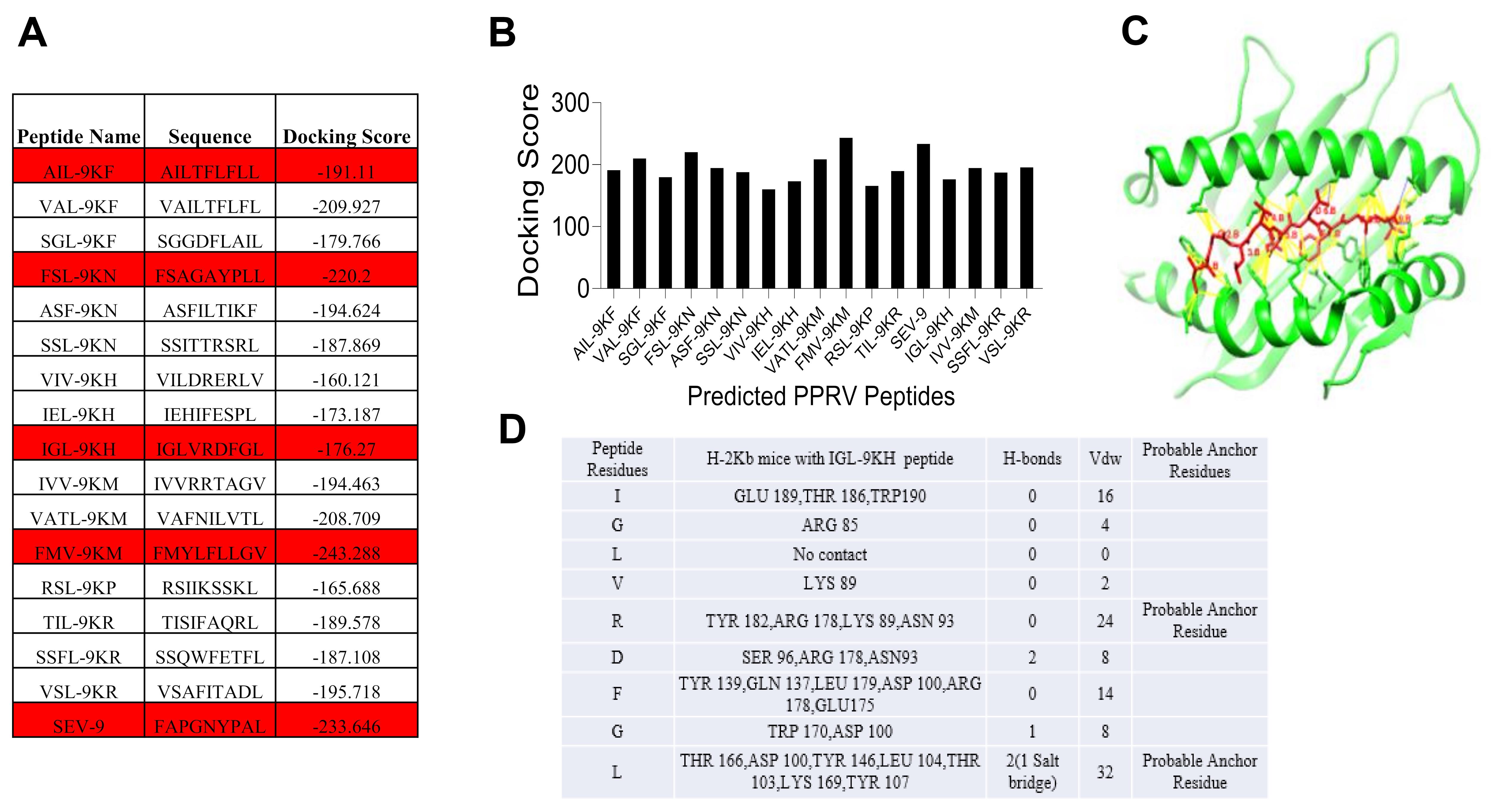
Figure 3. Molecular docking analysis of predicted class I MHC peptides with H-2Kb. Docking scores of the immunogenic predicted peptides are tabulated (A) and represented by a negative value of docking scores using a bar diagram (B). A docked image of a predicted peptide with the H-2Kb protein is shown (C). Different parameters and scores of individual residues in the peptide interacting with H-2Kb are tabulated (D). All the structures are reported in Sharma et al. (2021) .Generation of H-2Kb/peptide tetramers
To generate monomeric H-2Kb complexes, purified H-2Kb and β2m protein are refolded in the presence of high concentrations of H-2Kb restricted peptides. We have used a photo-cleavable peptide (FAPG[Anp]YPAL) to generate monomeric complexes, and biotinylated them to make multimers. A general scheme for refolding of a monomer and its tetramerization is shown in Figure 4. This photo-cleavable peptide can be easily exchanged with any designed H-2Kb restricted peptides by exposure to 365 nm UV wavelengths. A gel filtration chromatogram of a refolded monomer is shown (Figure 5). The lanes showing two distinct polypeptide bands corresponding to ~35 kDa and 13 kDa correspond to the units of the refolded monomers.
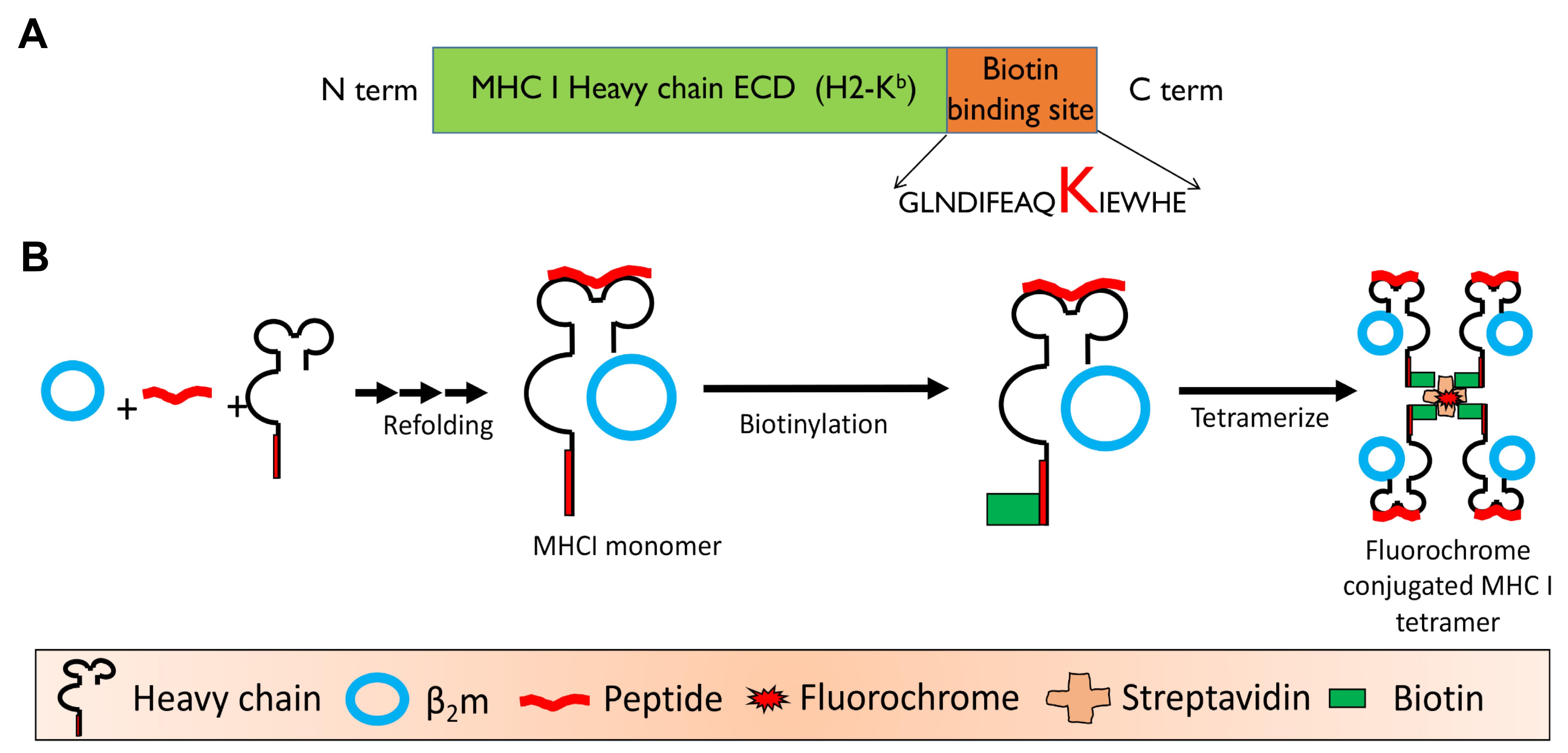
Figure 4. Schematic showing the generation of refolded heterotrimeric H-2Kb monomer consisting of peptide, β2m, and heavy chain of class I MHC (H-2Kb). A. Schematic of modified H-2Kb. B. Different steps in generating the class I MHC tetramers are depicted. The monomer was then biotinylated using a kit. The biotinylated monomers were tetramerized using a fluorescent avidin/streptavidin.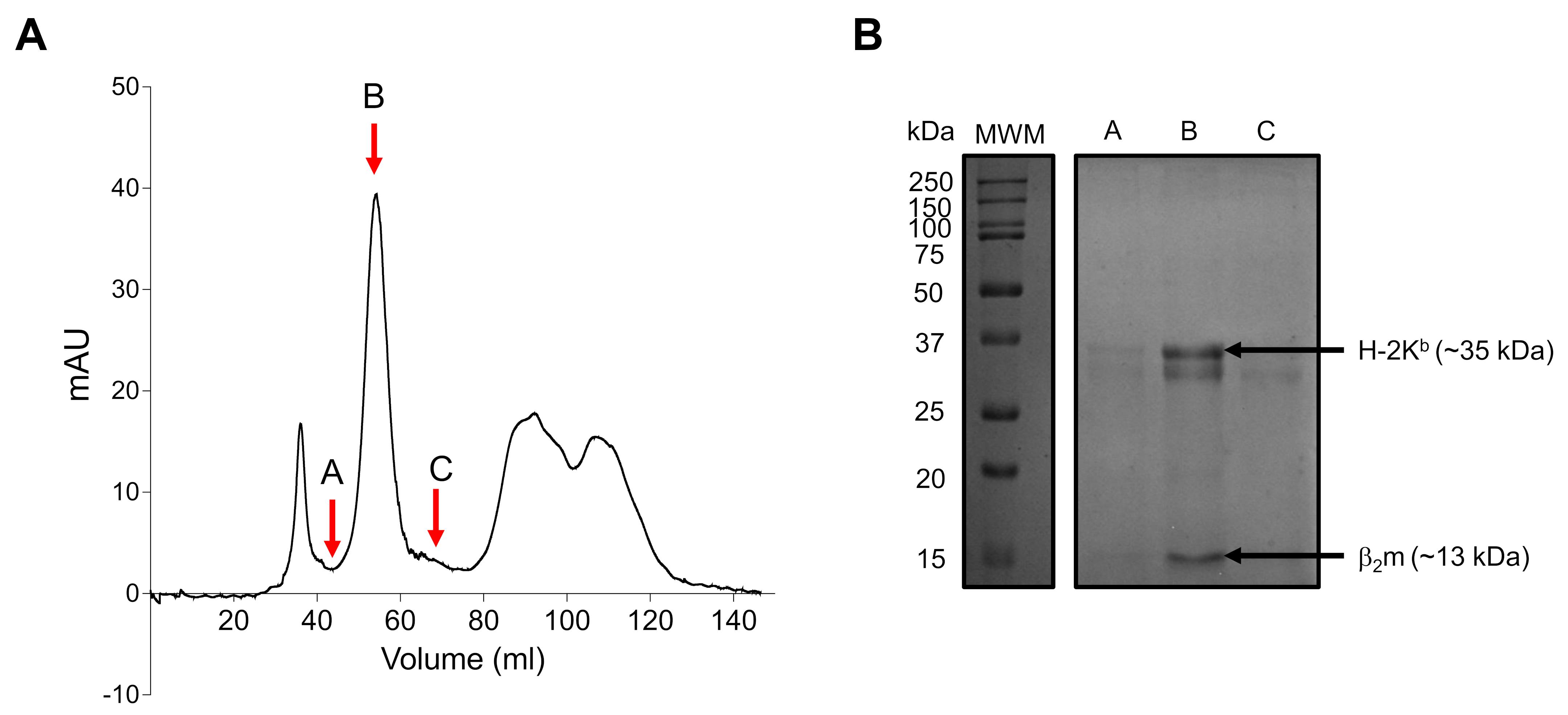
Figure 5. Refolding of H-2Kb monomers with conditional ligand. A. Chromatogram showing the elution profile of refolded H-2Kb monomers. The fractions marked A, B, and C were resolved using 12% SDA-PAGE (B). The presence of both H-2Kb heavy chain (≈35 kDa) and β2m (≈13 kDa) in fraction B shows a refolded monomer.Peptide/MHC I binding assay by acellular method
The purified monomers generated with a photocleavable peptide were used to select the best performing PPRV peptides in stabilizing plate bound class I MHC monomers following UV induced displacement of the conditional ligands. The stable monomer after peptide exchange will retain the β2m unit in the complex, and this can be easily detected with anti-β2m antibody using indirect ELISA; the lack of binding by the incoming peptide will disintegrate the complex, leading to loss of β2m detection (Figure 6A). The ELISA readings for all the peptides are compared with those of a known peptide (SIINFEKL) binding to H-2Kb, and are plotted as relative fold change with respect to the SIINFEKL peptide (Figure 6).
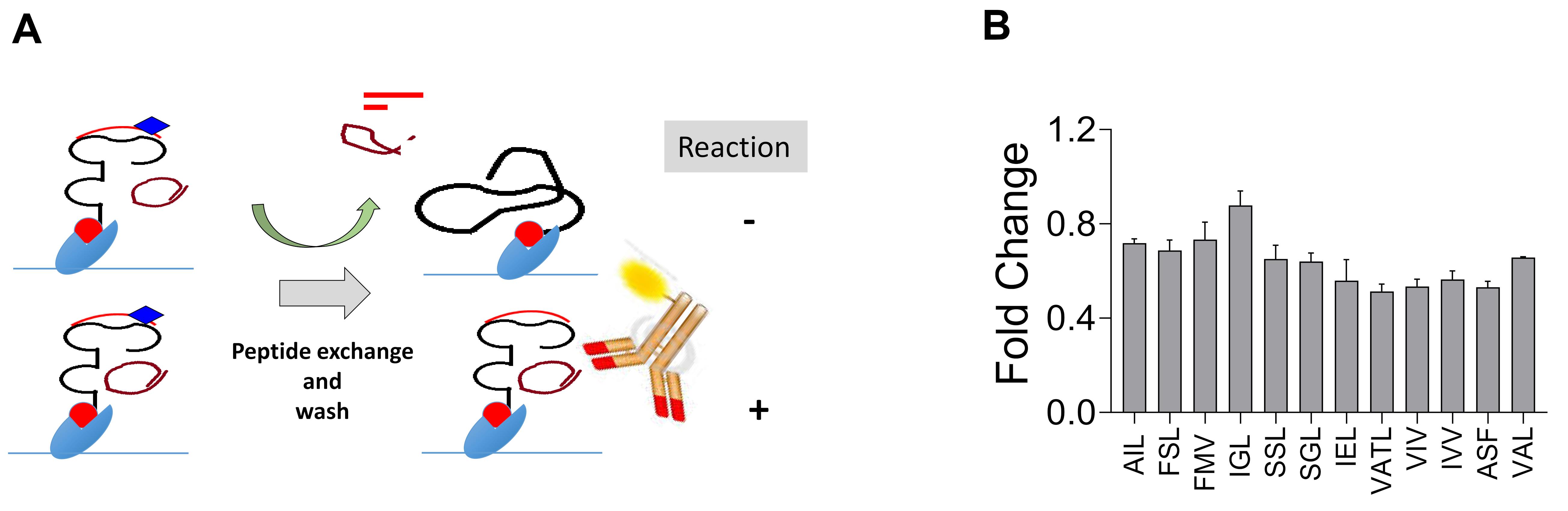
Figure 6. Acellular assay to determine peptide induced MHC stabilization. A. Schematic describing the binding of peptides with H-2Kb monomer with the conditional ligand after exposure to UV365. B. Bar diagram depicts fold change values of the binding affinity of different peptides of PPRV with H-2Kb following UV exchange compared to the positive control peptide (SIINFEKL). This data is reproduced from Sharma et al. (2021) . Only high scoring peptides are shown.Class I MHC stabilization by cellular method
To study the stabilization of class I MHC with different PPRV-peptides, we used TAP deficient RMA/S cell lines. RMA/S cells express very low levels of class I MHC I molecules on their cell surface. Since small peptides can be internalized by these cells, and get loaded on class I MHC molecule in yet to be adequately described subcellular compartments that also harbor class I MHC undergoing trafficking within the cell, to form a stable class I MHC complex. The peptide complexed class I MHC molecules are then expressed on the cell surface, and their expression levels directly correlate with the efficiency of peptide binding. The binding of exogenously added peptides with class I MHC could also occur on the cell surface. PPRV-peptides were exogenously added to the RMA/S cells, the expression of MHC I molecules was measured using anti-mouse H-2Kb antibody conjugated with fluorochrome, and detected using a flow cytometer (Figure 7).
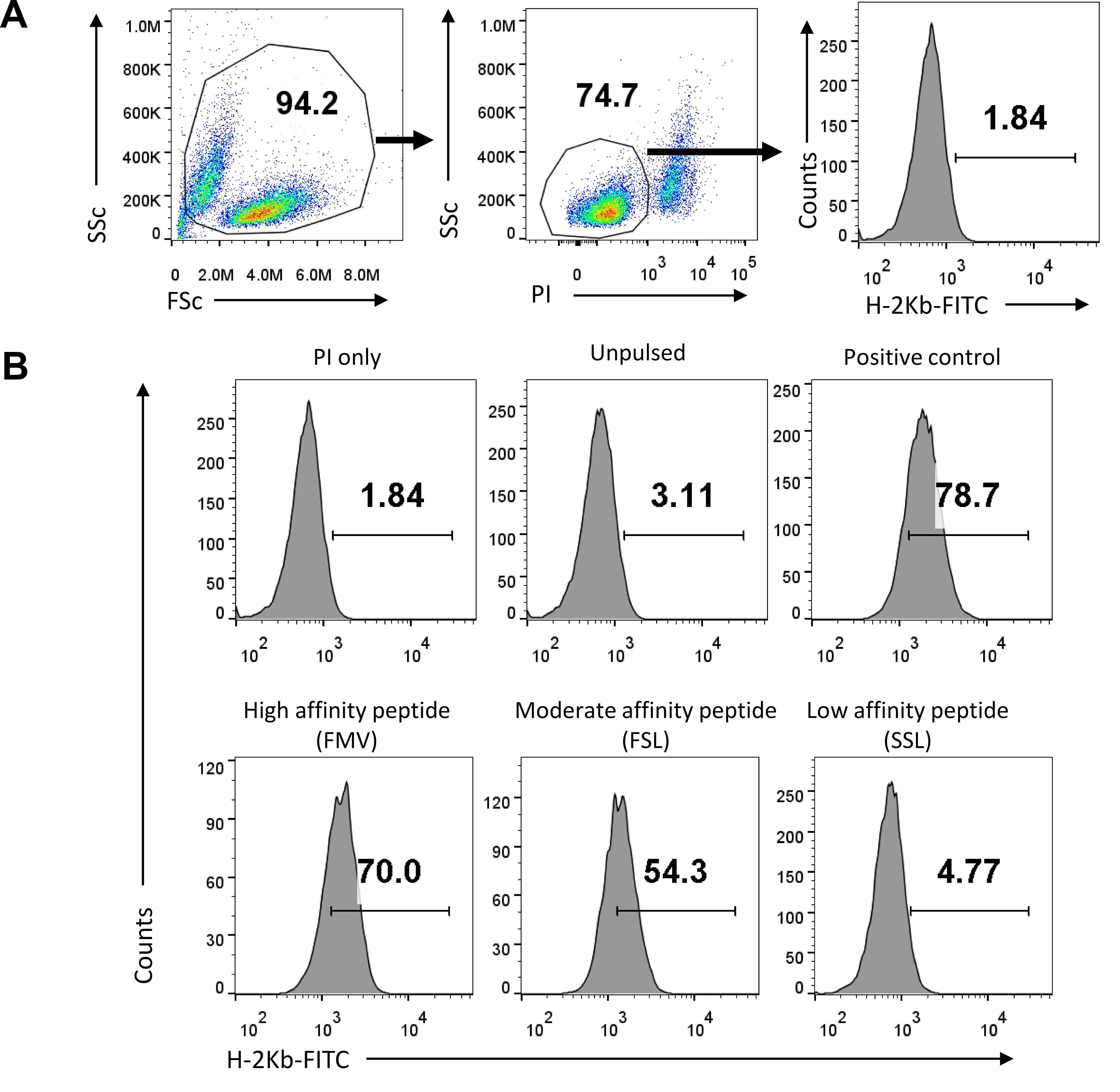
Figure 7. Flow cytometric analysis to quantify H-2Kb positive RMA/s cells. A. Representation of total cells gated using SSC and FSC plots. The gated cells were analyzed for propidium iodide (PI) staining and the PI negative live cells were measured for scoring H-2Kb positive staining. B. Representative flow histograms show the percentage of H-2Kb positive cells for high, moderate, and low affinity binding PPRV-peptides. SIINFEKL peptide was used as a positive control for MHC stabilization. Cells that had no added peptide, or those with a known peptide (SIINFEKL) were used as negative and positive controls, respectively. This data is reproduced from Sharma et al. (2021) .Detection of peptide specific CD8+ T cells using generated H-2Kb tetramers
The process of tetramerization of H-2Kb/peptide monomer involves the biotinylation of H-2Kb with biotin, and addition of streptavidin or avidin performed over a period of time. Four biotin binding sites in streptavidin generate H-2Kb/peptide tetramers. In order to use the tetramer for detection of peptide specific CD8+ T cells, monomers are tetramerized with fluorochrome conjugated streptavidin molecules. Blood samples from PPRV infected mice and PPRV peptide immunized mice were stained with anti-mouse CD8 antibody and the H-2Kb tetramer-APC. The analysis of peptide specific CD8+ T cells is shown in Figure 8, where double positive cells are gated to show the frequencies of PPRV-specific cells.
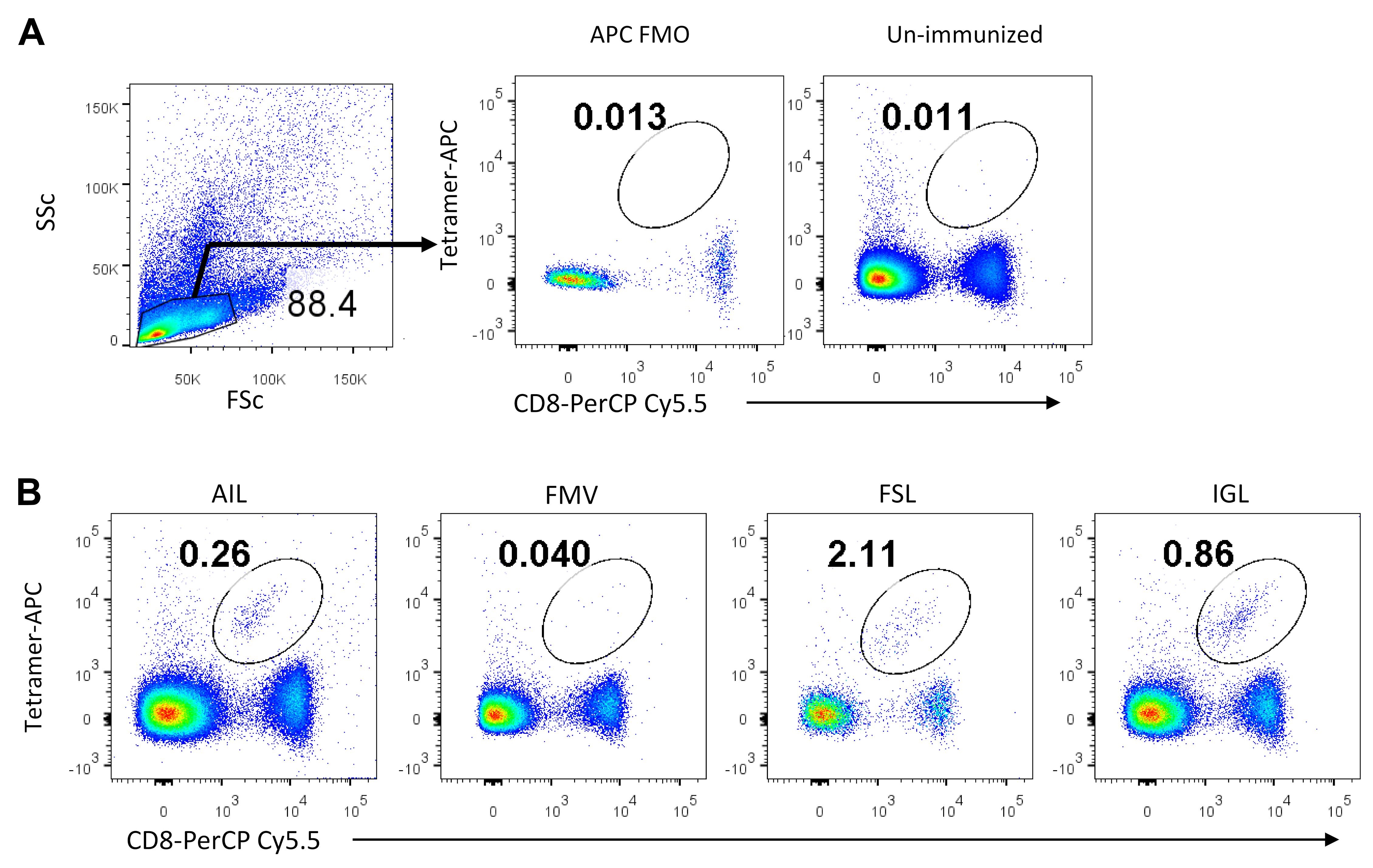
Figure 8. Flow cytometric analysis of peptide specific CD8+ T cells. A. PBMCs were first analysed using a SSC and FSC plot, and lymphocytes were gated. The gated population was then analysed for tetramer-APC on the Y-axis and CD8-PerCP Cy5.5 on the X-axis, using a bi-exponential FACS plot. Double positive cells (CD8+ Tetramer+) were gated to enumerate PPRV-peptide specific CD8+ T cells. B. Representative FACS plots show the frequencies of PPRV-specific CD8+ T cells after staining with tetramers generated by UV exchange process using different -PPRV peptides [AILTFLFLL (AIL), FMYLFLLGV (FMV), FSAGAYPLL (FSL) IGLVRDFGL (IGL), respectively]. Unexchanged tetramers served as the negative control for PPRV-specific cells. This data is reproduced from Sharma et al. (2021) .
Notes
Docking scores are based on the number of interactions the docked peptide shows with the class I MHC molecule. More interactions at desired sites lead to high docking scores (represented in negative values in Figure 2). Therefore, the higher the docking score, the better is the docking. Further scrutiny is done manually, based on additional parameters given in section B of the procedure.
Non-filtered tips were used for routine experiments, but for the cell culture experiments filtered tips were used.
During IPTG induction of recombinant proteins in bacteria, check OD600 appropriately. Do not proceed with induction if OD600 exceeds 0.8.
Before proceeding with the purification of recombinant proteins from inclusion bodies, check for the quality of induction of the recombinant proteins by resolving the uninduced and induced samples by SDS-PAGE.
While making the peptide emulsions for immunization, ensure proper synthesis by adding a small drop to the surface of ice cold water. A good emulsion preparation gives an intact drop for an extended duration.
The described protocol can be used to identify the immunogenic epitopes of any antigen derived from pathogens, cancers, or those responsible for autoimmune diseases.
Recipes
Stock solutions
Lysozyme
50 mg/ml in dH2O and store at -20°C
DNase I
2 mg/ml in 50% glycerol and 75 mM NaCl
MgCl2
0.5 M in dH2O and store at RT
Na·EDTA
0.5 M in 50 mM Tris-HCl, pH 8.0 and store at RT
Na·Azide
30% in PBS and store at RT
100 mM PMSF
174.2 mg in 10 ml 2-propanol and store at RT
1 M NaOH
Dissolve carefully 1 g of NaOH in 25 ml distilled water (dH2O).
LB media broth (1 L)
Tryptone 10 g
Yeast extract 5 g
Sodium chloride 10 g
Make up the volume to 1 L with dH2O.
Phosphate buffered saline, 1× PBS (for 1 L, pH 7.2-7.4)
137 mM NaCl 8 g
2.7 mM KCl 0.2 g
10 mM Na2HPO4 1.44 g
1.8 mM KH2PO4 0.24 g
Add 800 ml dH2O and adjust pH 7.2-7.4 and finally make up volume to 1 L with dH2O
PBS-T
0.05% Tween-20 in PBS (500 µl Tween in 1 L 1× PBS)
FACS Buffer
2% FBS in PBS (2 ml FBS in 100 ml 1× PBS)
Resuspension buffer (for 500 ml, pH 8.0)
50 mM Tris-HCl 3.94 g
25% Sucrose 125 g
1 mM Na·EDTA 186 mg
0.01% Na·Azide 1.6 ml (from 30% stock)
10 mM DTT 770 mg
1 mM PMSF 2 ml (from 100 mM stock)
Make up volume to 500 ml with dH2O
Lysis buffer (500 ml, pH 8.0)
50 mM Tris-HCl 3.94 g
1% Triton X-100 5 ml
100 mM NaCl 2.92 g
0.1% Na·Azide 1.6 ml (from 30% stock)
10 mM DTT 770 mg
Make up volume to 500 ml with dH2O
Washing buffer with Triton (for 500 ml, pH 8.0)
50 mM Tris-HCl 3.94 g
0.5% Triton X-100 2.5 ml
100 mM NaCl 2.93 g
1mM Na·EDTA 186 mg
0.1% Na·Azide 1.6 ml (from 30% stock)
1 mM DTT 77 mg
Make up volume to 500 ml with dH2O
Washing buffer without Triton (for 500 ml, pH 8.0)
50 mM Tris-HCl 3.94 g
100 mM NaCl 2.93 g
1mM Na·EDTA 186 mg
0.1% Na·Azide 1.6 ml (from 30% stock)
1 mM DTT 77 mg
Make up volume to 500 ml with dH2O
Urea buffer (for 500 ml)
8 M Urea 240.24 g
10 mM Na·EDTA 1.86 g
0.1 mM DTT 7.7 g
Make up volume to 500 ml with dH2O
Refolding buffer (for 250 ml, pH 8.0)
100 mM Tris-HCl 3.925 g
400 mM L-arginine-HCl 21 g
2 mM Na·EDTA 1 ml (from 0.5 M stock)
0.5 mM oxid. Glutathione 76.5 mg
5 mM red. Glutathione 3.75 g
Make up volume to 250 ml with dH2O
Guanidine solution (for 100 ml, pH 4.2)
3 M Guanidine-HCl 28.7 g
10 mM Na·Acetate 82 mg
10 mM Na·EDTA 2 ml (from 0.5 M stock)
Make up volume to 100 ml
Gel filtration buffer (for 500 ml, pH 8.0)
20 mM Tris-HCl 1.57 g
50 mM NaCl 1.46 g
Make up volume to 500 ml with dH2O and filter and de-gas
Red Blood Cell (RBC) Lysis Buffer (for 500 ml)
155 mM NH4Cl 4.14 g
12 mM NaHCO3 528 mg
0.1 mM EDTA 100 μl (from 0.5 M stock)
Make up volume upto 500 ml with dH2O
Acknowledgments
This protocol was primarily used in the research article Sharma et al. (2021). The authors would like to thank all the authors of Sharma et al. (2021) and Indian Institute of Science Education and Research Mohali and ICAR (NASF/ABA-6021/2017-2018 for providing funding to SS. The authors would like to thank Professor Hidde L Ploegh for providing the H-2Kb and β2m plasmid constructs. We also thank Surbhi Dahiya and Dhaneshwar Kumar of SS laboratory for the help provided during the preparation of the article.
Competing interests
The authors do not have any competing interests.
Ethics
The animal experiments used in all the experiments were performed strictly in accordance with the protocol (IISERM/SAFE/PRT/2018/014) approved by the Institutional Animal Ethics Committee (IAEC), Indian Institute of Science Education and Research Mohali, constituted under the aegis of committee for the purpose of control and supervision of experiments on animals (CPCSEA).
References
- Chen, J. E., Huang, C. C. and Ferrin, T. E. (2015). RRDistMaps: a UCSF Chimera tool for viewing and comparing protein distance maps. Bioinformatics 31(9): 1484-1486.
- De Bruijn, M. L., Schumacher, T. N., Nieland, J. D., Ploegh, H. L., Kast, W. M. and Melief, C. J. (1991). Peptide loading of empty major histocompatibility complex molecules on RMA-S cells allows the induction of primary cytotoxic T lymphocyte responses. Eur J Immunol 21(12): 2963-2970.
- De Silva, A. D., Boesteanu, A., Song, R., Nagy, N., Harhaj, E., Harding, C. V. and Joyce, S. (1999). Thermolabile H-2Kb molecules expressed by transporter associated with antigen processing-deficient RMA-S cells are occupied by low-affinity peptides. J Immunol 163(8): 4413-4420.
- Esquivel, F., Yewdell, J. and Bennink, J. (1992). RMA/S cells present endogenously synthesized cytosolic proteins to class I-restricted cytotoxic T lymphocytes. J Exp Med 175(1): 163-168.
- Garboczi, D. N., Hung, D. T. and Wiley, D. C. (1992). HLA-A2-peptide complexes: refolding and crystallization of molecules expressed in Escherichia coli and complexed with single antigenic peptides. Proc Natl Acad Sci U S A 89(8): 3429-3433.
- Garstka, M. A., Fish, A., Celie, P. H., Joosten, R. P., Janssen, G. M., Berlin, I., Hoppes, R., Stadnik, M., Janssen, L., Ovaa, H., et al. (2015). The first step of peptide selection in antigen presentation by MHC class I molecules. Proc Natl Acad Sci U S A 112(5): 1505-1510.
- La Gruta, N. L., Gras, S., Daley, S. R., Thomas, P. G. and Rossjohn, J. (2018). Understanding the drivers of MHC restriction of T cell receptors. Nat Rev Immunol 18(7): 467-478.
- Pettersen, E. F., Goddard, T. D., Huang, C. C., Couch, G. S., Greenblatt, D. M., Meng, E. C. and Ferrin, T. E. (2004). UCSF Chimera--a visualization system for exploratory research and analysis. J Comput Chem 25(13): 1605-1612.
- Pichugin, A., Zarling, S., Perazzo, L., Duffy, P. E., Ploegh, H. L. and Krzych, U. (2018). Identification of a Novel CD8 T Cell Epitope Derived from Plasmodium berghei Protective Liver-Stage Antigen. Front Immunol 9: 91.
- Princiotta, M. F., Finzi, D., Qian, S. B., Gibbs, J., Schuchmann, S., Buttgereit, F., Bennink, J. R. and Yewdell, J. W. (2003). Quantitating protein synthesis, degradation, and endogenous antigen processing. Immunity 18(3): 343-354.
- Rodenko, B., Toebes, M., Hadrup, S. R., van Esch, W. J., Molenaar, A. M., Schumacher, T. N. and Ovaa, H. (2006). Generation of peptide-MHC class I complexes through UV-mediated ligand exchange. Nat Protoc 1(3): 1120-1132.
- Schumacher, T. N., Heemels, M. T., Neefjes, J. J., Kast, W. M., Melief, C. J. and Ploegh, H. L. (1990). Direct binding of peptide to empty MHC class I molecules on intact cells and in vitro. Cell 62(3): 563-567.
- Sehrawat, S., Kirak, O., Koenig, P. A., Isaacson, M. K., Marques, S., Bozkurt, G., Simas, J. P., Jaenisch, R. and Ploegh, H. L. (2012). CD8+ T cells from mice transnuclear for a TCR that recognizes a single H-2Kb-restricted MHV68 epitope derived from gB-ORF8 help control infection. Cell Rep 1(5): 461-471.
- Sharma, Y., Sarkar, R., Jain, A., Singh, S., Shekhar, C., Shanmugam, C., Dhanavelu, M., Tembhurne, P., Kaul, R. and Sehrawat, S. (2021). A Mouse Model of PPRV Infection for Elucidating Protective and Pathological Roles of Immune Cells. Front Immunol 12: 630307.
- Vita, R., Mahajan, S., Overton, J. A., Dhanda, S. K., Martini, S., Cantrell, J. R., Wheeler, D. K., Sette, A. and Peters, B. (2019). The Immune Epitope Database (IEDB): 2018 update. Nucleic Acids Res 47(D1): D339-D343.
- Vita, R., Overton, J. A., Greenbaum, J. A., Ponomarenko, J., Clark, J. D., Cantrell, J. R., Wheeler, D. K., Gabbard, J. L., Hix, D., Sette, A. et al. (2015). The immune epitope database (IEDB) 3.0. Nucleic Acids Res 43(Database issue): D405-412.
- Vyas, J. M., Van der Veen, A. G. and Ploegh, H. L. (2008). The known unknowns of antigen processing and presentation. Nat Rev Immunol 8(8): 607-618.
- Wieczorek, M., Abualrous, E. T., Sticht, J., Alvaro-Benito, M., Stolzenberg, S., Noe, F. and Freund, C. (2017). Major Histocompatibility Complex (MHC) Class I and MHC Class II Proteins: Conformational Plasticity in Antigen Presentation. Front Immunol 8: 292.
- Xu, T., Li, X., Wu, Y., Shahzad, K. A., Wang, W., Zhang, L. and Shen, C. (2016). A Combination Strategy for Construction of Peptide-β2m-H-2Kb Single Chain with Overlap Extension PCR and One-Step Cloning. J Microbiol Biotechnol 26(12): 2184-2191.
- Zhang, X. W. (2013). A combination of epitope prediction and molecular docking allows for good identification of MHC class I restricted T-cell epitopes. Comput Biol Chem 45: 30-35.
- Zhou, P., Jin, B., Li, H. and Huang, S. Y. (2018). HPEPDOCK: a web server for blind peptide-protein docking based on a hierarchical algorithm. Nucleic Acids Res 46(W1): W443-W450.
Article Information
Copyright
© 2021 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Sarkar, R., Sharma, Y., Jain, A., Tehseen, A., Singh, S. and Sehrawat, S. (2021). A Combinatorial in-silico, in-vitro and in-vivo Approach to Quantitatively Study Peptide Induced MHC Stability. Bio-protocol 11(24): e4255. DOI: 10.21769/BioProtoc.4255.
Category
Immunology > Immune cell staining > Flow cytometry
Immunology > Animal model > Mouse
Biological Sciences > Biological techniques
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.