- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Isolation and Culturing Primary Cholangiocytes from Mouse Liver

Published: Vol 11, Iss 20, Oct 20, 2021 DOI: 10.21769/BioProtoc.4192 Views: 5946
Reviewed by: Masahiro MoritaKomuraiah Myakala
Original research article
The authors used this protocol in:

Jul 2021

Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Related protocols
Abstract
Cholangiocytes are epithelial cells lining the intrahepatic and extrahepatic bile ducts. Cholangiocytes perform key physiological functions in the liver. Bile synthesized by hepatocytes is secreted into bile canaliculi, further stored in the gallbladder, and finally discharged into the duodenum. Due to liver injury, biliary epithelial proliferate in response to endogenous or exogenous signals leading to cholangiopathies, inflammation, fibrosis, and cholangiocarcinoma. Cholangiocytes exhibit anatomical and functional heterogeneity, and understanding such diversified functions will potentially help in finding effective therapies for various cholestatic liver diseases. To perform such functional studies, effective cholangiocyte isolation and culture procedures are needed. This protocol will aid in easy isolation and expansion of cholangiocytes from the liver.
Keywords: CholangiocytesBackground
The liver is structurally and functionally heterogeneous. It performs vital functions like the uptake of cholesterol; amino acid, lipid, and vitamin metabolism; and the production of albumin and blood clotting factors. The liver is composed of parenchymal and non-parenchymal cells, of which hepatocytes form a major portion; other cells present include biliary epithelial cells (cholangiocytes), stellate cells, liver sinusoidal endothelial cells, and various immune cell subsets (Zorn, 2008). Liver sinusoidal endothelial cells form a lining between the blood in the sinusoidal space and underlying hepatocytes, forming the space of Disse. Liver lobules are functional units that have a central vein from which hepatocyte cords arise and are separated into sinusoids carrying blood from the portal triad to the central vein. Portal triads are composed of the hepatic artery, bile duct, and portal vein. Bile ducts are lined by epithelial cells called cholangiocytes (Malarkey et al., 2005; Gordillo et al., 2015), as shown in Figure 1. The biliary epithelium primarily acts as a lining conduit for bile flow, but it also modifies canalicular bile and concentrates bile in the gall bladder. Biliary epithelia are “effective communicators” with neighboring cells in producing mediators that are involved in cell growth and injury response (Chen et al., 2008). Many risk factors, including genetic mutations, immune-mediated damage, idiopathic factors, infectious agents including viruses, and vascular defects, can modify cholangiocyte physiology, resulting in altered bile transport leading to cholestasis, which further induces inflammation, fibrosis, and end-stage liver cirrhosis (Alvaro et al., 2007; Sebode et al., 2014; Asai et al., 2015; Miethke et al., 2016; Carey et al., 2017; Pinto et al., 2018; Taylor et al., 2018; Banales et al., 2019; Pham et al., 2021).

Figure 1. Architecture of the liver. The liver is composed of functional units called lobules. Lobules consist of portal triads containing a bile duct, hepatic artery, and portal vein. Lobules have hepatic cords with a number of sinusoids. Cholangiocytes form a lining around the bile ducts. Adapted from Gordillo et al. (2015).
Disorders of the extrahepatic bile duct cause increased susceptibility to morbidity and mortality. Biliary atresia, primary sclerosing cholangitis, cystic fibrosis, Alagille’s syndrome, and primary biliary cirrhosis are some of the well documented cholangiopathies affecting both neonates and elderly patients. About 70% of pediatric liver transplants are performed to treat biliary atresia, while 5% of liver transplantations are done in patients with primary sclerosing cholangitis (PSC) (Kelly and Davenport, 2007; Wunsch et al., 2014; Govindarajan, 2016; Lazaridis and LaRusso, 2016; Fickert and Wagner, 2017; Dyson et al., 2018; Shah et al., 2020). However, the exact etiology or the factors that mediate biliary injury in some patients are still unknown. The susceptibility of these patients can be better understood using murine models and cholangiocytes isolated from these models through in vitro experiments. However, the culture of primary biliary epithelial cells remains challenging. This protocol will help facilitate the isolation and expansion of native cholangiocytes, which can be further used for downstream applications.
Materials and Reagents
100 µm nylon mesh (Greiner Bio-One, catalog number: 542000)
50 ml syringe (BD, Catalog number: 309653)
50 ml tubes (Greiner Bio-One, catalog number: 227270)
15 ml tubes (Greiner Bio-One, catalog number: 188261)
5 ml pipette (Greiner Bio-One, catalog number: 606180)
48-well plates (Fisher Scientific, catalog number: 08-772-1C)
T-25 and T-75 flasks (Greiner Bio-One, catalog numbers: 1186Q51 and 658175)
Pasteur pipet (Fisher, catalog number: 1367820D)
Dexamethasone (400 μg/ml) (Sigma, catalog number: D4902)
3,3’,5-Triiodo-L-thyronine (3.4 mg/ml) (Sigma, catalog number: T6397)
Forskolin (0.411 mg/ml) (Sigma, catalog number: F3917)
mEGF (10μg/ml) (Peprotech, catalog number: 315-09)
ITS (100×) (Corning Cellgro ITS, catalog number: MT25800CR)
L-glutamine (100×) (Gibco Life Technologies, catalog number: 102374)
Pen/Strep (100×) (Thermo Fisher Scientific, catalog number: 15070063)
MEM-NEAA (100×) (Gibco, catalog number: 11140050)
EpCam G8.8 Antibody (University of Iowa Hybridoma bank)
Percoll (GE Healthcare, catalog number: 17089101)
PBS (Fisher, catalog number: 14190250)
Hyaluronidase (Sigma-Aldrich, catalog number: H3506)
Collagenase D (Sigma-Aldrich, catalog number: 11088882001)
EpCam (murine) (CD326, G8.8; DSHB, https://dshb.biology.uiowa.edu/G8-8)
Sheep Anti-Rat IgG Dynabeads (Thermo Fisher Scientific, catalog number: 11035)
DMEM w/ F12 (Gibco, catalog number: 11320033)
MEM-NEAA (Gibco, catalog number: 11140050)
Triiodothyronine (Sigma-Aldrich, catalog number: T6397)
PierceTM BCA Protein Assay Kit (Thermo Fisher Scientific, catalog number: 23227)
Rat anti-mouse CK19 IgG, Troma III (CK19) (DSHB, https://dshb.biology.uiowa.edu/TROMA-III)
Goat anti Rat antibody (Invitrogen, Fisher Scientific, catalog number: PI31470)
Donkey anti-Mouse IgG Secondary Antibody IRDye 800CW LI-COR (Biosciences, catalog number: 926-32212)
DNase I (Roche, catalog number: 4716728001)
Release Buffer (Thermo Fisher Scientific)
EasySepTM Release Mouse PE Positive Selection Kit (Stem Cell Technology, catalog number: 17656)
PE anti-mouse CD326 (Ep-CAM), Antibody (Biolegend, catalog number: 118206)
DMEM w/ F1 (Gibco, catalog number: 11320033)
FBS (Thermo Fisher Scientific, catalog number: A4766801)
Chemically defined Lipid Concentrate (100×) (Gibco, catalog number: 11905031)
Gentamicin (50 mg/ml) (Gibco, catalog number: 15750-060)
MEM Vitamin Solution (100×) (Gibco, catalog number: 11120052)
Glacial acetic acid (Fisher Scientific, catalog number: A38S-500)
Pierce BCA Protein Assay Kit (Thermo Scientific, catalog number: PI23235)
β-actin (Sigma-Aldrich, catalog number: A5441)
Pierce Fast Western Blot Kit, ECL Substrate (Thermo Scientific, catalog number: 35055)
Donkey anti-mouse IgG IRDye 800CW (LI-COR Biosciences, catalog number: 926-32212)
Horseradish peroxidase (Fisher Scientific, catalog number: PI31470)
Odyssey system (LI-COR) (https://www.licor.com/bio/odyssey-clx/)
DPBS (Fisher, catalog number: 14190 250)
Rat Tail Collagen Coating Solution (20 ml) (Cell Applications, catalog number: 122-20)
Media for digestion/Perfusion
Perfusing media (see Recipes)
Digestion media (see Recipes)
Isolation buffer (see Recipes)
Dexamethasone (400 μg/ml) (see Recipes)
3,3’,5-Triiodo-L-thyronine (3.4 mg/ml) (see Recipes)
Forskolin (0.411 mg/ml) (see Recipes)
mEGF (10 μg/ml) (see Recipes)
Collagen coating (see Recipes)
Equipment
Hemocytometer (Sigma, catalog number: Z359629-1EA)
Centrifuge (Eppendorf, catalog number: EP-5810S)
Rotator (Cambridge Scientific, catalog number: 17847)
Forceps and scalpel (Fine Science Tools, catalog number: 11075-00; VWR, catalog number: 4-422)
Shaker (Eppendorf, catalog number: M1352-0000)
BD Accuri C6 flow cytometer (BD Biosciences)
BD LSRFortessa Cell Analyzer (BD Biosciences, https://www.bdbiosciences.com/en-in/products/instruments/flow-cytometers/research-cell-analyzers/bd-lsrfortessa)
Bio-Rad ChemiDocTM MP Imaging System
Software
FlowJo software (version 10.7.1; Tree Star, Inc., Ashland, OR)
Procedure
Harvest and tissue dissociation
Anesthetize mice using isoflurane.
Open the abdominal cavity. Using 50 ml syringe, perfuse the liver with 30 ml of perfusing media through the inferior vena cava with a flow rate of 5 ml/min, and cut the portal vein to drain the perfusion media.
Remove the liver and weigh it to correlate cell number with liver weight.
Scrap the perfused liver with a scalpel; this will help to separate the bile ducts from the rest of the liver parenchyma. Mince the bile duct and left-over liver tissue mass into very small pieces using forceps and a scalpel.
Place the minced pieces in 50 ml tubes containing 10 ml of digestion media.
Place the 50 ml tubes in the shaker and shake at 280 rpm for 30 min at 37°C.
After 30 min of constant shaking, remove the tubes from the shaker and add 5 ml (33%) of heat-inactivated FBS. The addition of FBS deactivates enzymatic activity.
Pass the cell suspension through a 100 µm nylon mesh cell strainer into a 50 ml tube.
Use a 3 ml syringe plunger to scrape and press tissue pieces through the strainer. Wash the strainer with 35 ml of isolation buffer and make up the volume to 50 ml.
Centrifuge at 300 × g for 7 min at 4°C.
Wash the cells two times with isolation buffer.
Cell isolation by gradient centrifugation
Make percoll gradient solutions as shown in Table 1.
Table 1. Concentration of percoll gradient solutions used for cell isolation
Density Vol. of percoll PBS Final vol. 26.25% 6.3 ml 17.7 ml 24 ml 41.5% 6.65 ml 9.35 ml 16 ml 73.3% 8.8 ml 3.2 ml 12 ml The calculations shown in Table 1 are sufficient for four livers.
Place a Pasteur pipet in the 15 ml tube.
Resuspend the cells in 6 ml of 26.25% density gradient percoll and add into 15 ml tube with Pasteur pipet.
Using a 5 ml pipette, add 4 ml of 41.5% density percoll through the Pasteur pipet and let it flow under the 26.25% gradient layer in the tube. Add 3 ml of 73.3% percoll solution through the Pasteur pipet in the tube. The 73.3% density solution will layer under the 41.5% density solution, as shown in Figure 2.
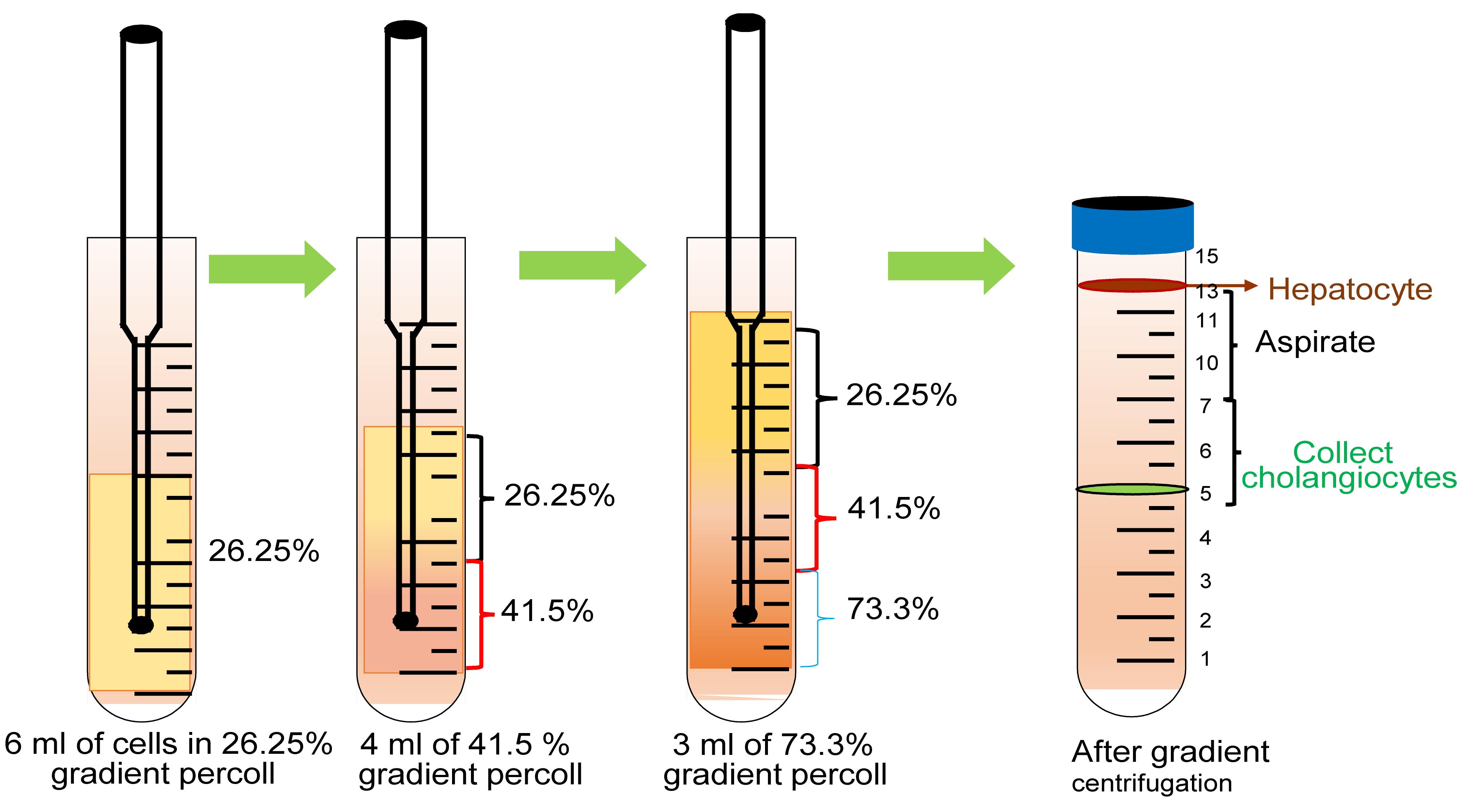
Figure 2. Overlaying of percoll gradient solutionCentrifuge at 796 × g for 30 min at 4°C without breaks.
Remove the 15 ml tubes from the centrifuge.
Aspirate the upper layer of hepatocytes and collect to 3 ml of interphase containing cholangiocytes; place it in the 50 ml tube.
Wash the cells using 50 ml of isolation buffer by centrifuging at 300 × g for 7 min at 4°C. Repeat another time; this will help remove the percoll.
Remove the supernatant and resuspend the cell pellet in 1 ml of isolation buffer.
Assess the fraction of Epcam positive cells using the Epcam PeCy7 labeled flow antibody on a BD Accuri C6 flow cytometer and BD LSRFortessa Cell Analyzer (Figure 3).
Count the cells using a hemocytometer.
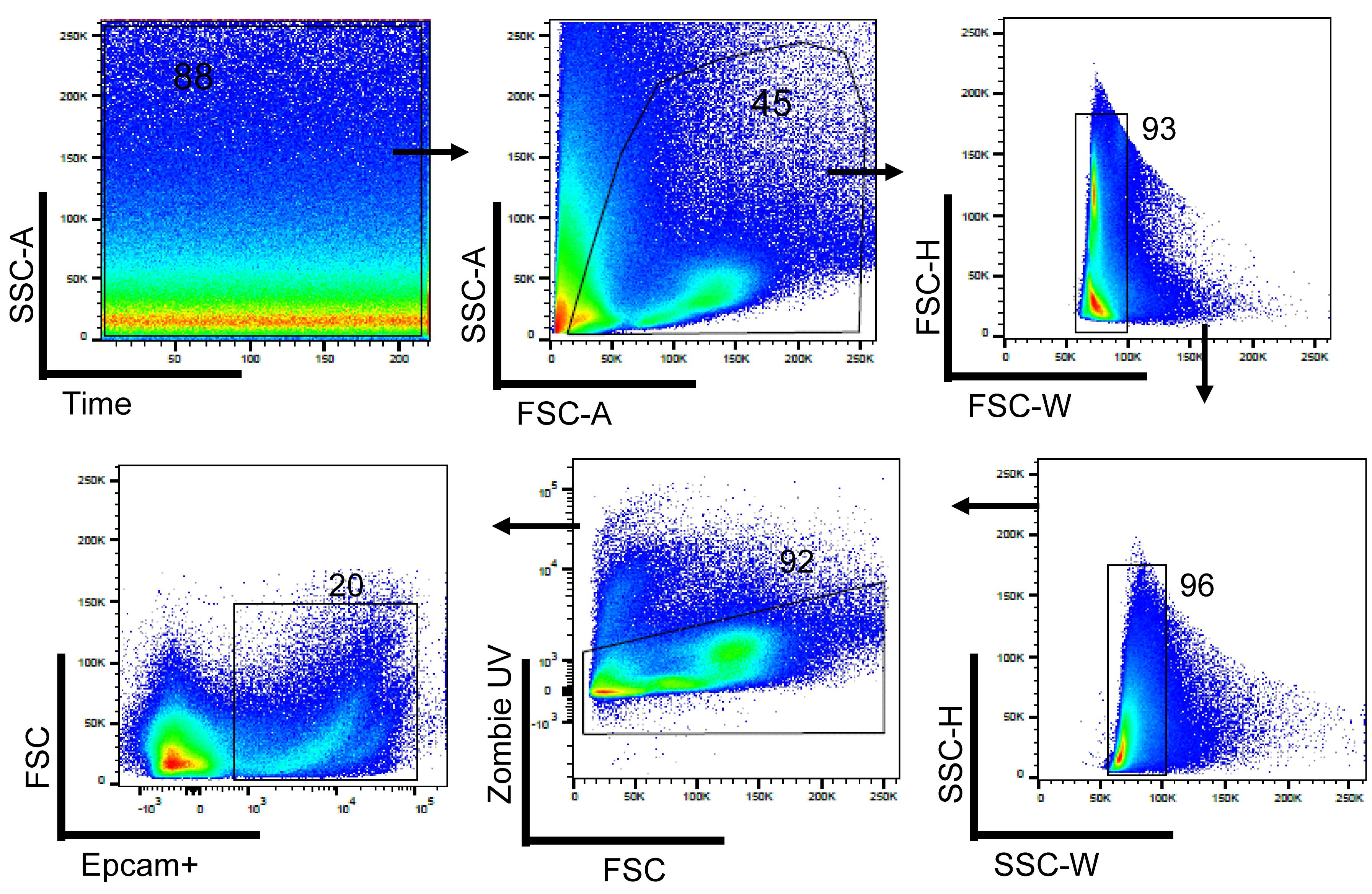
Figure 3. Fraction of Epcam+ cholangiocytes isolated with the gradient centrifugation method
Purification/enrichment of cholangiocytes using the Dynabead cell isolation kit
Wash the Dynabeads before proceeding to cell isolation.
Add desired volume (based on cell number) of beads to a tube. Wash the Dynabeads using 1 ml of isolation buffer.
Place the tube in a magnet for 2 min and discard the supernatant.
Remove the 15 ml tube from the magnet and resuspend the beads in the 1 ml of isolation buffer.
Use approximately 10 μg of EpCAM G8.8 primary antibody (rat IgG) per 107 cells, mix well, and incubate at 4°C on a rotator for 20 min.
Wash the cells by adding 2 ml of isolation buffer and centrifuge at 300 × g for 7 min. Discard the supernatant.
Resuspend the cells in isolation buffer. Add 20 μl of anti-rat IgG Dynabeads to 15 ml tube with cells. Incubate at 4°C on a rotator for 20 min.
After incubating with Dynabeads, add 1 ml of isolation buffer, pipet up and down 2-3 times, and place the tube in a magnet for 4 min.
For positive isolation, while the tube is still in the magnet, carefully remove and discard the supernatant containing unbound cells. Repeat Step C8 one more time to remove unbound cells; this will increase the purity of isolated cells.
Resuspend the cell pellet in isolation buffer and wash the cells by centrifuging at 300 × g for 7 min at 4°C. Remove the supernatant and proceed.
Releasing bead-bound cells
Resuspend the bead-bound cells in 1 ml of release buffer.
Incubate for 15 min with rolling and tilting at room temperature, and then mix by pipetting up and down 10 times, preventing the formation of air bubbles. This will help in efficiently separating the cells from beads.
Place the tube in a magnet for 4 min.
Transfer the supernatant containing the bead-free cholangiocytes into a new 15 ml tube.
Add Isolation buffer to 15 ml and centrifuge at 300 × g for 7 min. Discard the supernatant and resuspend the cell pellet in 1ml of cholangiocyte culture media. Count the cells using a hemocytometer.
Keep the cells from 2°C to 8°C until further use in downstream applications.
Cells isolated from neonatal cholangiocytes are expanded as described below.
Note: In addition to the above-discussed method. Epcam positive cells can be purified using the PE anti-mouse CD326 (Ep-CAM) Antibody and EasySepTM Release Mouse PE Positive Selection Kit. This method can be used after counting cells with a hemocytometer in Procedure B.
Expansion of cholangiocytes
Media: To make a 500 ml stock, add the following reagents in the appropriate concentrations (Table 2).
Table 2. List of reagents used to prepare 500 ml of stock media
Reagents Volume 1 DMEM w/ F12 450 ml 2 FBS 50 ml 3 L-glutamine (100×) 5 ml 4 Pen/Strep (100×) 5 ml 5 MEM-NEAA (100×) 5 ml 6 Chemically defined
Lipid Concentrate (100×)5 ml 7 Gentamicin (50 mg/ml) 1 ml MEM Vitamin Solution (100×) 5 ml Note: To make complete media, add the following supplements in Table 3 on the day staring cell culture.
Table 3. List of reagents added to stock to make complete media for the expansion of cholangiocytes
Reagents Volume 1 DMEM w/ F12 with supplement (Table 2) 50 ml 2 ITS (100×) 500 µl 3 Dexamethasone (400 μg/ml) 50 µl 4 Triiodothyronine (3.4 mg/ml) 50 µl 5 mEGF (10 μg/ml) 125 µl 6 Forskolin (0.411 mg/ml) 50 µl Note: The complete media containing ITS, dexamethasone, triiodothyronine, mEGF, and Forskolin is usable for 1 week. Gentamicin (100 µg/ml) is used for cholangiocyte culture.
Culturing/Passaging
Note: For initial cultures, start with 48-well plates and move to 12-well plate once cells are confluent; then, transfer the cells to T-25 or T-75 flasks depending on requirements. Cell growth is very slow for the first few passages.
T-25 flask
To coat 1 T-25 flask, add 31 µl of collagen (4 mg/ml) + 3 ml of 0.1% acetic acid
T-75 flask
To coat 1 T-75, add 94 µl of collagen (4 mg/ml) + 5 ml of 0.1% acetic acid
Incubate for 2 h at room temperature or overnight at 4°C.
Remove coating solution by washing with 5 ml of sterile PBS (when using T-25 flask).
Gently rock the flask with PBS; repeat washing three to four times to remove acetic acid.
If passaging, dissociate the adhered cells using TrypLE. Prewarm TrypLE to 37°C.
Wash cells with 5 ml of DPBS without Ca++ and Mg++. Aspirate and discard.
Add an appropriate volume (2 ml in T-25, 2-5 ml in T-75 flask) of TrypLE to flask.
Incubate at 37°C for 2 to 4 min until cells detach. Gently tap the flask to dislodge cells if necessary.
Wash cells with 13 ml DPBS without Ca++ and Mg++. Transfer the cell suspension to a 15 ml conical tube.
Centrifuge at 300 × g for 7 min 4°C. Discard the supernatant and resuspend the cell pellet in pre-warmed complete medium. Count the cells and divide them accordingly. After a few passages, cells grow fast; therefore, split the cells based on requirements.
Check the purity of cultured cells using Epcam PeCy7 labeled flow antibody on the BD Accuri C6 flow cytometer (Figure 4).
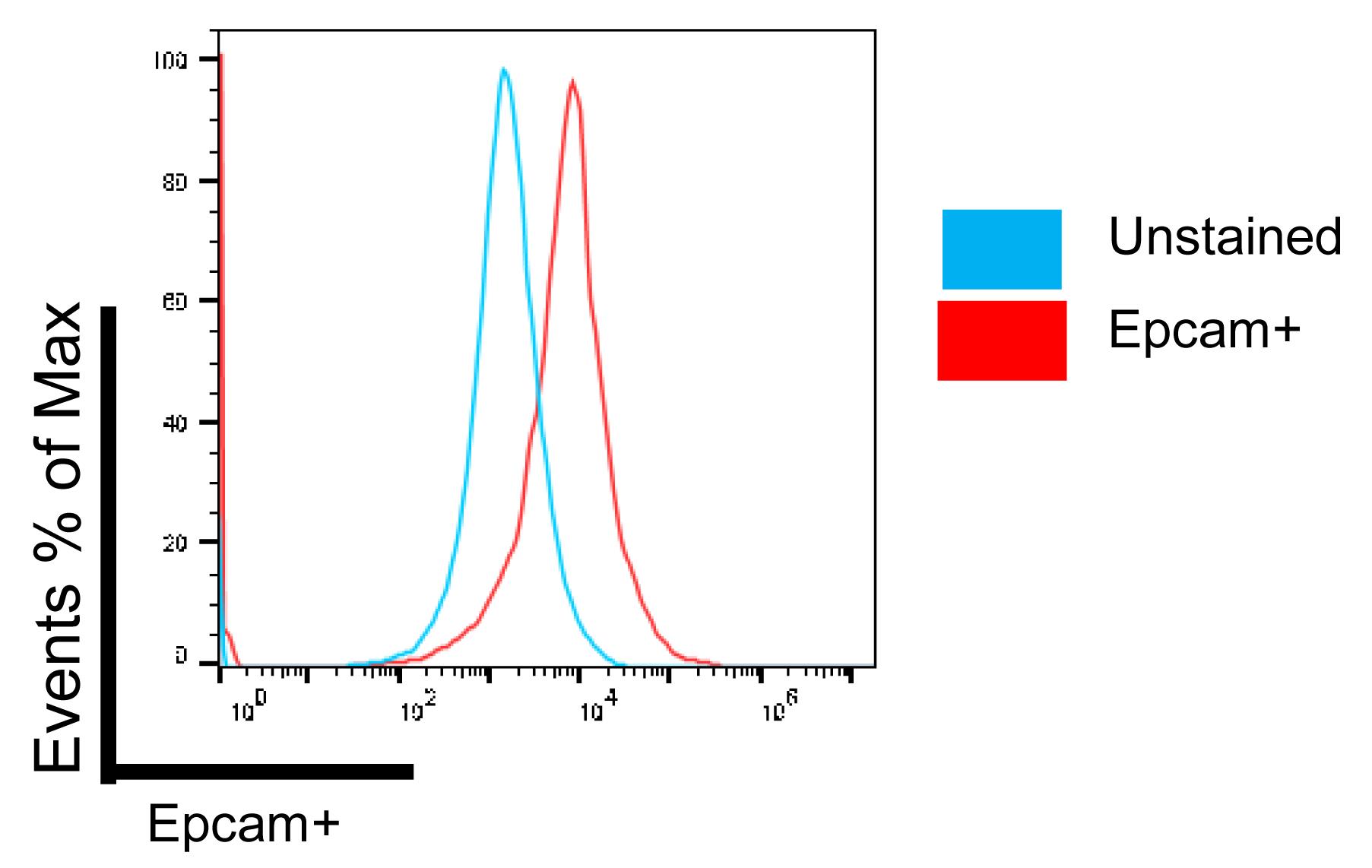
Figure 4. Fraction of Epcam+ cholangiocytes from passage 6Immunoblotting for CK19 and β-actin was performed on purified and in vitro expanded cholangiocytes. Protein was quantified using the PierceTM BCA Protein Assay Kit, and 35 μg of protein per lane was used.
For detection of β-actin and CK19, membranes were incubated at 4°C overnight with primary antibody.
On the next day, blots were washed and incubated with horseradish peroxidase secondary antibody. CK19 was detected using a Bio-Rad ChemiDocTM MP Imaging System using the ECL Substrate.
β-actin, used as internal control, was detected with donkey anti-mouse IgG secondary antibody. Immunoblots were visualized using the Odyssey system (LI-COR) (Figure 5).
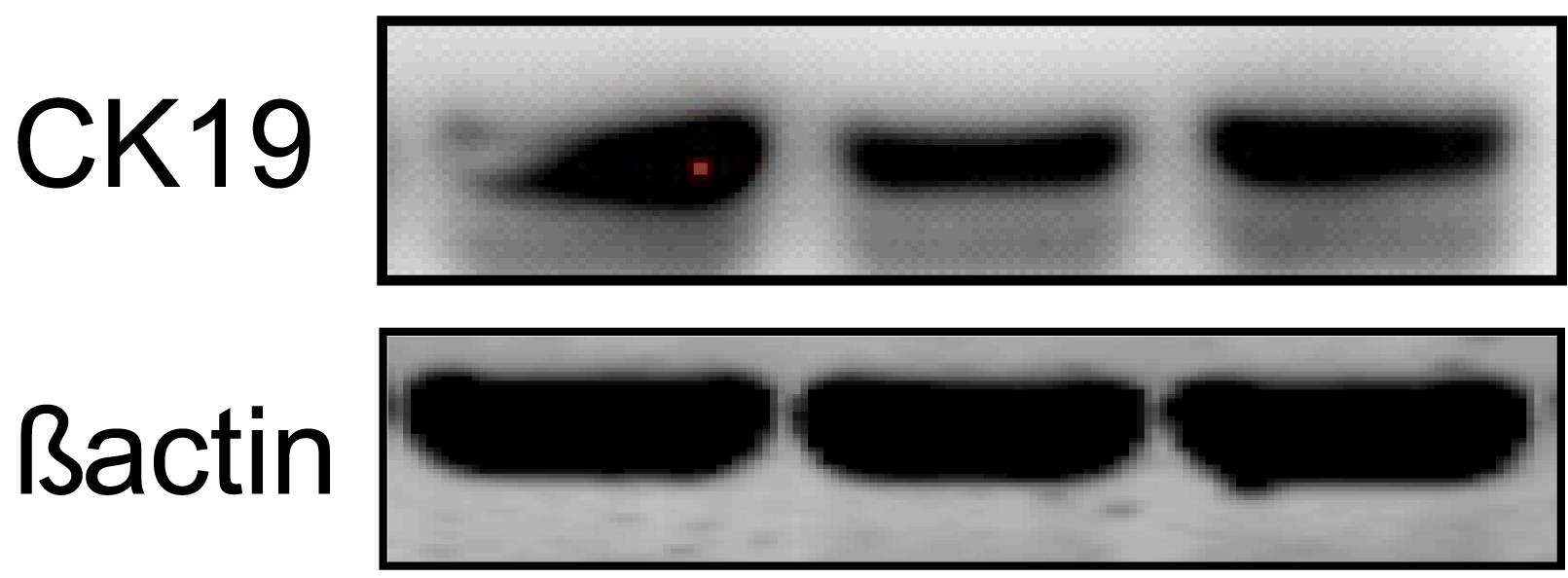
Figure 5. CK19 expression in cholangiocytes from passage 6
Summary
Cholestasis is defined as an impairment in bile formation or flow, while the etiology of cholestasis differs, ranging from genetic mutation to autoimmunity. Bile is the digestive fluid produced in the liver by hepatocytes, and disorders in hepatocytes, biliary ducts/biliary cells, and pancreas can cause cholestasis causing retention of bile salts in the body. Bile acids precipitated during cholestasis mediate biliary injury and further cause obliteration of cholangiocytes, a cellular lining for bile ducts known as the biliary tree. Cholangiocyte damage causes obstruction of bile transport and bile modification. We identified a homozygous, nonsense pathogenic variant in ABCC12 encoding an adenosine 5′-triphosphate (ATP)-binding cassette (ABC)-transporter, multidrug resistance-associated protein 9 (MRP9) in a patient. These patients elicit bile duct paucity while showing lesser GGT levels. Expression of MRP9 transporters was restricted to cholangiocytes in the liver. So far, the role of this transporter has not been discussed in the liver. Therefore, we aimed to explore the role of ABCC12 and its biological function using primary cholangiocytes isolated from neonates and adult mice. Biliary epithelial cells were isolated from 2-day-old pups and from adult mice. In adult mice, the liver was perfused with DMEM+F12 media containing collagenase D. Liver tissue was diced into small pieces and further digested with Collagenase D, Hyaluronidase, and DNase I. Then, 5 ml of heat-inactivated FBS was added to digested tissue to inactivate enzymes. The digest was further washed with DMEM+F12 containing 10% FBS. Three different concentrations/densities of percoll were used to isolate cholangiocytes. Cells resuspended in percoll were centrifugated at 2,000 rpm for 30 min without brakes. The interface layer was collected, washed with DMEM+F12, and incubated with the EpCAM Ab to purify cholangiocytes from the rest of the liver cells. Cells were washed and incubated with Sheep Anti-Rat IgG Dynabeads, and the bead-bound Epcam positive cells were separated from other cells using magnetic columns. Epcam positive cells were used for different experiments or expanded in vitro to identify the role of ABCC12 in cholangiocytes. This protocol was used in our recent study published in Gastroenterology (Pham et al., 2021).
Recipes
Perfusing media
50 ml of DMEM F12 + 500 µl (1 mg/ml) of Collagenase D + 1% FBS
Digestion media
30 ml DMEM + 15 mg Collagenase D + 15 mg Hyaluronidase + 200 μl of 200 mM CaCl2 + 200 µl of 200 mM MgSO4, DNases I (10 U/ml).
Isolation Buffer
500 ml of Ca2+ and Mg2+ free PBS supplemented with 0.1% BSA and 2 ml of 0.5 M; final concentration is 2 mM EDTA and pH 7.4.
Dexamethasone (400 μg/ml)
Weigh 4 mg of dexamethasone.
Use 2-4 ml of ethanol to dissolve.
Add sterile media to 10 ml.
3,3’,5-Triiodo-L-thyronine (3.4 mg/ml)
Dissolve 100 mg of Triiodo-L-thyronine powder in 2 ml of 1 N NaOH.
Place in 50 ml tube.
Add 27.4 ml of sterile media for a total of 29.4 ml.
Forskolin (0.411 mg/ml)
Dissolve 10 mg of Forskolin in 5% DMSO/H2O (1.2 ml DMSO + 23.1 ml sterile H2O).
mEGF (10 μg/ml)
Dissolve 100 µg in 10 ml 1× PBS/1% BSA.
Collagen coating 4 mg/ml
Make a 0.1% (~0.02 M) acetic acid solution: dilute glacial acetic acid (17.5 M) 1:1,000 in dH2O. Calculate amount of collagen to coat dishes at 5 µg/cm2. To coat one T-25 flask, add 31 µl of collagen (4 mg/ml) to 3 ml of 0.1% acetic acid.
Acknowledgments
The authors would like to acknowledge support for this work from the Center for Autoimmune Liver Disease at Cincinnati Children’s Hospital Medical Center and R01 DK095001 (A.M.).
Competing interests
All authors declare no conflict of interest.
Ethics
The Laboratory animal care and use committee of the Cincinnati Children’s Research Foundation (Cincinnati, OH) approved the work described here.
References
- Alvaro, D., Mancino, M. G., Glaser, S., Gaudio, E., Marzioni, M., Francis, H. and Alpini, G. (2007). Proliferating cholangiocytes: a neuroendocrine compartment in the diseased liver. Gastroenterology 132(1): 415-431.
- Asai, A., Miethke, A. and Bezerra, J. A. (2015). Pathogenesis of biliary atresia: defining biology to understand clinical phenotypes. Nat Rev Gastroenterol Hepatol 12(6): 342-352.
- Banales, J. M., Huebert, R. C., Karlsen, T., Strazzabosco, M., LaRusso, N. F. and Gores, G. J. (2019). Cholangiocyte pathobiology. Nat Rev Gastroenterol Hepatol 16(5): 269-281.
- Carey, A. N., Zhang, W., Setchell, K. D. R., Simmons, J. R., Shi, T., Lages, C. S., Mullen, M., Carroll, K., Karns, R., Bessho, K., et al. (2017). Hepatic MDR3 expression impacts lipid homeostasis and susceptibility to inflammatory bile duct obstruction in neonates. Pediatr Res 82(1): 122-132.
- Chen, X. M., O'Hara, S. P. and LaRusso, N. F. (2008). The immunobiology of cholangiocytes. Immunol Cell Biol 86(6): 497-505.
- Dyson, J. K., Beuers, U., Jones, D. E. J., Lohse, A. W. and Hudson, M. (2018). Primary sclerosing cholangitis. Lancet 391(10139): 2547-2559.
- Fickert, P. and Wagner, M. (2017). Biliary bile acids in hepatobiliary injury - What is the link? J Hepatol 67(3): 619-631.
- Gordillo, M., Evans, T. and Gouon-Evans, V. (2015). Orchestrating liver development. Development 142(12): 2094-2108.
- Govindarajan, K. K. (2016). Biliary atresia: Where do we stand now? World J Hepatol 8(36): 1593-1601.
- Kelly, D. A. and Davenport, M. (2007). Current management of biliary atresia. Arch Dis Child 92(12): 1132-1135.
- Lazaridis, K. N. and LaRusso, N. F. (2016). Primary Sclerosing Cholangitis. N Engl J Med 375(25): 2501-2502.
- Malarkey, D. E., Johnson, K., Ryan, L., Boorman, G. and Maronpot, R. R. (2005). New insights into functional aspects of liver morphology. Toxicol Pathol 33(1): 27-34.
- Miethke, A. G., Zhang, W., Simmons, J., Taylor, A. E., Shi, T., Shanmukhappa, S. K., Karns, R., White, S., Jegga, A. G., Lages, C. S., et al. (2016). Pharmacological inhibition of apical sodium-dependent bile acid transporter changes bile composition and blocks progression of sclerosing cholangitis in multidrug resistance 2 knockout mice. Hepatology 63(2): 512-523.
- Pham, D. H., Kudira, R., Xu, L., Valencia, C. A., Ellis, J. L., Shi, T., Evason, K. J., Osuji, I., Matuschek, N., Pfuhler, L., et al. (2021). Deleterious Variants in ABCC12 are Detected in Idiopathic Chronic Cholestasis and Cause Intrahepatic Bile Duct Loss in Model Organisms. Gastroenterology 161(1): 287-300 e216.
- Pinto, C., Giordano, D. M., Maroni, L. and Marzioni, M. (2018). Role of inflammation and proinflammatory cytokines in cholangiocyte pathophysiology. Biochim Biophys Acta Mol Basis Dis 1864(4 Pt B): 1270-1278.
- Sebode, M., Peiseler, M., Franke, B., Schwinge, D., Schoknecht, T., Wortmann, F., Quaas, A., Petersen, B. S., Ellinghaus, E., Baron, U., et al. (2014). Reduced FOXP3+ regulatory T cells in patients with primary sclerosing cholangitis are associated with IL2RA gene polymorphisms. J Hepatol 60(5): 1010-1016.
- Shah, A., Macdonald, G. A., Morrison, M. and Holtmann, G. (2020). Targeting the Gut Microbiome as a Treatment for Primary Sclerosing Cholangitis: A Conceptional Framework. Am J Gastroenterol 115(6): 814-822.
- Taylor, A. E., Carey, A. N., Kudira, R., Lages, C. S., Shi, T., Lam, S., Karns, R., Simmons, J., Shanmukhappa, K., Almanan, M., et al. (2018). Interleukin 2 Promotes Hepatic Regulatory T Cell Responses and Protects From Biliary Fibrosis in Murine Sclerosing Cholangitis. Hepatology 68(5): 1905-1921.
- Wunsch, E., Trottier, J., Milkiewicz, M., Raszeja-Wyszomirska, J., Hirschfield, G. M., Barbier, O. and Milkiewicz, P. (2014). Prospective evaluation of ursodeoxycholic acid withdrawal in patients with primary sclerosing cholangitis. Hepatology 60(3): 931-940.
- Zorn, A. (2008). Liver development. In: StemBook (Internet). Cambridge, MA: Harvard Stem Cell Institute.
Article Information
Copyright
© 2021 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Kudira, R., Sharma, B. K., Mullen, M., Mohanty, S. K., Donnelly, B., Tiao, G. M. and Miethke, A. (2021). Isolation and Culturing Primary Cholangiocytes from Mouse Liver. Bio-protocol 11(20): e4192. DOI: 10.21769/BioProtoc.4192.
Category
Developmental Biology > Cell growth and fate > Regeneration
Immunology > Animal model > Mouse
Cell Biology > Cell isolation and culture > Cell isolation > Flow cytometry
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.