- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

In situ Hybridization of miRNAs in Human Embryonic Kidney and Human Pluripotent Stem Cell-derived Kidney Organoids

Published: Vol 11, Iss 17, Sep 5, 2021 DOI: 10.21769/BioProtoc.4150 Views: 3359
Reviewed by: Giusy TornilloAnonymous reviewer(s)
Original research article
The authors used this protocol in:
Jan 2021
Advertisement


Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Related protocols
Abstract
MicroRNAs are small RNAs that negatively regulate gene expression and play an important role in fine-tuning molecular pathways during development. There is increasing interest in studying their function in the kidney, but the majority of studies to date use kidney cell lines and assess the total amounts of miRNAs of interest either by qPCR or by high-throughput methods such as next generation sequencing. However, this provides little information as to the distribution of the miRNAs in the developing kidney, which is crucial in deciphering their role, especially as there are multiple kidney cell types, each with its own specific transcriptome. Thus, we present a protocol for obtaining spatial information for miRNA expression during kidney development by in situ hybridization (ISH) of anti-miRNA, digoxigenin-labelled (DIG), Locked Nucleic Acid (LNA®) probes on (i) native human embryonic tissue and (ii) human pluripotent stem cell (hPSC)-derived 3D kidney organoids that model kidney development. We found that the method reveals the precise localization of miRNA in specific anatomical structures and/or cell types and confirms their absence from others, thus informing as to their specific role during development.
Keywords: Kidney developmentBackground
MicroRNAs (miRNAs) are small (20-25 nucleotides) RNAs that regulate gene expression by binding predominantly to the 3’ UTRs of their target genes’ mRNAs and inhibiting their translation and/or causing their degradation (Bartel, 2018). An increasing number of studies now show that miRNAs play a crucial role in fine-tuning molecular pathways in the kidney, both in normal development and in disease (Jones et al., 2018; Trionfini et al., 2015; Zhao et al., 2019). The majority of these studies have been conducted on animal models, which do not always faithfully recapitulate the developmental events or disease phenotypes in humans. To overcome these limitations, we recently reported using human pluripotent stem cell (hPSC)-derived kidney organoids (Bantounas et al., 2018; Takasato et al., 2015) as a model to study the role of the miR-199a/214 cluster during human kidney development (Bantounas et al., 2021). In that study, we used in situ hybridization of miRNAs with digoxigenin-labelled locked-nucleic acid (LNA®) probes to detect the precise localization of the miRNAs of this cluster in paraffin sections of both native kidney and organoids. LNA® probes have a chemically modified backbone to afford a higher melting temperature (Tm) per nucleotide than conventional DNA or RNA probes (Singh et al., 1998). They are, thus, ideally suited to the detection of smaller RNAs for which conventional probes would have too low a Tm to bind efficiently. After binding to the tissue, the probes are detected by application of an alkaline phosphatase (AP)-linked, anti-digoxigenin antibody, followed by the addition of an AP substrate, which is converted to a coloured product, visible under a light microscope. Using this method, we discovered that miR-199a-3p and miR-214-3p were both present in the kidney stroma and in developing glomeruli but were largely absent from mature glomeruli. One of the miRNAs, miR-214-3p, also exhibited strong tubular expression. In addition, we observed differences in the extent and distribution of expression between embryonic/fetal kidney and organoids (particularly in the case of miR-199a-3p), possibly indicating that the two represented different developmental stages (Bantounas et al., 2021). This example demonstrates the main advantage of this method, which is the acquisition of spatial information regarding the expression of the miRNAs of interest within a given tissue. This information would be lost if one were to rely exclusively on transcriptional detection methods [e.g., qPCR or Next Generation miRNA Sequencing (miRseq)]. The hybridization steps of the protocol presented here (see below) were adapted from Jørgensen et al. (2010) and were optimized for kidney/kidney organoids and the particular miRNAs we studied (Bantounas et al., 2021). However, the method can in principle be used with formalin-fixed, paraffin-embedded (FFPE) sections of any tissue and for any miRNA or other small RNA of similar size.
Materials and Reagents
1.5 ml Eppendorf tubes (Starlab, catalog number: S1615-5550)
Plastic pipette tips (preferably filtered) (Starlab, catalog numbers: S1122-1830 [1,000 μl]; S1120-8810 [200 μl]; S1123-1810 [20 μl]; S1121-3810 [10 μl]; or equivalent)
Vacuum filter units, pore size 0.2 μm (Thermo, catalog number: 568-0020)
6-well plates (Costar, catalog number: 3516, or equivalent)
3 ml plastic Pasteur pipettes (Starlab, catalog number: E1414-0311, or equivalent)
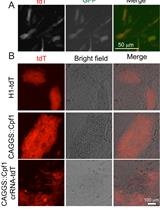
Tissue processing/embedding cassettes (Simport, catalog number: M490-4) (Figure 1, Left)
Stainless steel or plastic tissue processing capsules (Figure 1, Right)
Note: The capsules depicted in Figure 1 were the ones used by the authors but have now been discontinued. Therefore, alternative options are proposed here: Simport, catalog number: M470; or Fisher, catalog number: 15-182-219; or equivalent.

Figure 1. Tissue processing cassette and capsuleMetal molds for paraffin embedding of tissue (Leica, catalog number: 3803081E)
RNaseZap® wipes for disinfection of working surfaces (Invitrogen/Ambion, catalog number: AM9788)
Superfrost® Plus slides (ThermoFisher Scientific, catalog number: J1800AMNZ)
Human embryonic kidney tissue was provided by the MRC and Wellcome Trust Human Developmental Biology Resource (http://www.hdbr.org/)
hPSC-derived kidney organoids can be produced according to published protocols (Takasato et al., 2015; Bantounas et al., 2018 and 2021). In our study (Bantounas et al., 2021), we differentiated the MAN13 human embryonic stem cell (hESC) line (Ye et al., 2017), but any hESC or induced pluripotent stem cell (iPSC) line can be used in principle
Paraffin wax for embedding tissue, melting point 61°C (Pfm Medical, catalog number: 9000)
Nuclease-free water (not DEPC-treated) (ThermoFisher Scientific, catalog number: AM9930)
Phosphate buffered saline (PBS), without Ca2+ and Mg2+ (Sigma, catalog number: D8537-500 ml)
5’-Digoxigenin-labelled miRCURY® LNA® miRNA detection probes (1 nmol) (see Note 1) (QIAGEN, catalog number: 339111; see below for individual probe codes). In this example, we used probes targeting:
miR-199a-3p (QIAGEN, catalog number: YD00615410)
miR-214-3p (QIAGEN, catalog number: YD00611471)
scrambled probe (negative control; QIAGEN, catalog number: YD00699004)
U6 snRNA (positive/optimization control; QIAGEN, catalog number: YD00699002)
miRCURY® LNA® miRNA ISH Buffer Set (FFPE) (QIAGEN, catalog number: 339450), which includes:
2× Formamide-free miRNA ISH buffer
Proteinase K solution
UltraPureTM 20× SSC Buffer (Invitrogen, catalog number: 15557-044)
Tween-20 (Sigma, catalog number: P1379-250ML)
Sheep anti-DIG-AP (alkaline phosphatase linked) antibody (Roche/Sigma-Aldrich, catalog number: 11093274910)
Sheep serum (Sigma-Aldrich, catalog number: S3772-10ML)
Bovine serum albumin (BSA) (Sigma-Aldrich, catalog number: A1470-100G)
Nitro blue tetrazolium/5-bromo-4-chloro-indolyl-phosphate (NBT/BCIP) ready-to-use tablets (Roche, catalog number: 1 1 697 471 001)
Levamisol (Fluka, catalog number: 31742), diluted in water to make a 100 mM stock solution
Nuclear Fast Red solution (Merck, catalog number: N3020)
Industrial methylated spirit (IMS) (various suppliers)
Xylene (various suppliers)
100% Ethanol (various suppliers)
KCl (various suppliers)
KP Marker Plus (Histolab, catalog number: 98307-R) or similar hydrophobic marking pen (various suppliers)
4% Paraformaldehyde (PFA) Solution (see Recipes)
1 M Tris Buffer (see Recipes)
0.5 M EDTA Solution (see Recipes)
5 M NaCl Solution (see Recipes)
1× Proteinase K Buffer (see Recipes)
SSC Buffer dilutions (see Recipes)
PBS-T 0.1% (see Recipes)
Blocking solution (see Recipes)
Antibody dilution buffer (see Recipes)
KTBT Buffer (see Recipes)
Equipment
Tissue Processor (Leica, model: ASP300S; or equivalent)
Heated Paraffin Embedding Station (Leica HistoCore Arcadia H or equivalent)
Cold Plate (Leica, HistoCore Arcadia C or equivalent)
Microtome (Leica, model: RM225; or equivalent)
Hybridization oven or other variable temperature incubator (e.g., Hybaid, Midi Dual 14 or equivalent)
Microscope slide rack (Simport, catalog number: M905-12DGY)
Jars for immersing the slide rack into solvents/buffers (Simport, catalog number: M900-12G)
Heating block (up to 90°C) (Eppendorf ThermoMixer F1.5 or equivalent)
Microcentrifuge (Labnet, Prism or equivalent)
Gilson® or equivalent pipettes (various suppliers)
Staining tray for holding the slides during hybridization and staining (see Figure 2).
Note: The tray depicted in Figure 2 was the one used by the authors but has now been discontinued. An alternative is proposed here: Fisher, catalog number: 22-045-035.
Waterbath (Grant SUB6, catalog number: P266; or equivalent)
Autoclave (various suppliers)
Optionally, for imaging: 3D-Histech Panoramic-250 microscope slide-scanner, with a 40×/0.95 Plan Apochromat objective (Zeiss)
Software
Optional: CaseViewer (3DHISTECH Ltd.; www.3dhistech.com), to capture images following scanning with a 3D-Histech slide-scanner (see point 14, in Equipment)
Fiji/ImageJ (http://imagej.net/Fiji/Downloads) (for image processing following capture)
Procedure
Tissue Fixation
The procedure described in this section is for preparing organoid tissue and is an adaptation of the one described in Lopes et al. (2019).
If beginning with samples already embedded in paraffin: Skip directly to section B, step 10.
The procedure below assumes organoids cultured in transwell inserts in 6-well plates (Bantounas et al., 2018). If a different size well and/or insert is used, change volumes proportionally in Steps A1-A7.
Prepare “wash” and “fixation” 6-well plates, allowing one well per transwell insert of organoids:
Wash plate(s): Add 1.2 ml PBS into each well.
Fixation plate(s): Add 1.2 ml of 4% PFA into each well.
Transfer the transwell inserts from the culture plate to the wash plate prepared above.
Carefully add 1 ml of PBS into each transwell and swirl the plate gently to wash the organoids.
Using a Gilson or a 3 ml Pasteur pipette, remove the PBS from inside the transwells.
Transfer the transwells from the “wash” to the “fixation” plate (i.e., on top of the PFA that is already in each of the wells).
Add 1 ml 4% PFA into each transwell making sure that the organoids are fully covered.
Note: You may add more PFA, if necessary, to completely cover the organoids.
Incubate at room temperature for 20 min.
Remove the PFA from the transwells and transfer them to a new wash plate, prepared as in Step A1a, above.
Wash the organoids twice with PBS by repeating Steps A2-A4 as above.
Using a 3 ml plastic Pasteur pipette, take some of the PBS of the last wash in the transwell and expel it onto each organoid, with just enough force so as to dislodge it (but take care not to be too forceful, which can break up the organoid).
Using the same 3 ml Pasteur, transfer each organoid into a separate fresh 1.5 ml Eppendorf tube containing 400 μl PBS. The fixed organoids can be stored at 4°C in PBS for up to one week before embedding.
Embedding fixed tissue in paraffin and sectioning
A visual guide to the embedding and sectioning of tissues, which is also applicable to our organoids in this section, can be found at this address: https://www.youtube.com/watch?v=7-LIbAWPc-g (accessed in 2021).
Label a number of plastic tissue embedding cassettes (point 8, in Materials and Reagents; Figure 1, Left) equal to the number of organoids with a pencil or permanent marker pen to distinguish between samples.
Place a tissue processing capsule (point 9, in Materials and Reagents; Figure 1, Right) into each plastic tissue cassette, both with lids off.
Using a plastic Pasteur pipette, carefully transfer one organoid into the capsule base.
Replace the lid on the tissue processing capsule, followed by the lid on the tissue cassette.
Immerse the cassette into 70% ethanol.
Repeat Steps B3-B5 for each organoid.
Transfer the cassettes to the tissue processor (Leica ASP300S or equivalent) and run a program following the steps as listed in Table 1.
Table 1. Tissue processing program (R.T.: room temperature)
The next day, take the cassettes out of the tissue processor and transfer them into paraffin wax, using a Heated Paraffin Embedding Station, as follows:
Open the plastic cassette followed by the tissue processing capsule.
With the aid of liquid wax from the embedding station, carefully push the organoid into a metal mold.
Carefully position the organoid in the middle of the mold.
Allow to set for a few seconds on a Cold Plate (see Equipment). Top up the mold with liquid wax, place the plastic cassette, without lid, on the top of the mold and leave it to solidify on the cold plate for about 2 h.
Repeat Steps B8a-B8d for each organoid.
Place the blocks in a container of melting ice/icy water for at least 15 min before sectioning and using a microtome, produce serial 5 µm sections of the paraffin block containing the organoid.
Allow the tissue sections to extend (so that any creases are straightened out) on the surface of clean warm tap water and then transfer them to labelled microscope slides. Allow to dry out on a warm plate and transfer to a 37°C oven overnight for further drying.
Step/Solvent Time Temperature 70% IMS 20 min R.T. 70% IMS 30 min R.T. 90% IMS 45 min R.T. 90%IMS 60 min R.T. 100% Absolute Alcohol 30 min R.T. 100% Absolute Alcohol 45 min R.T. 100% Absolute Alcohol 60 min R.T. Xylene 20 min 40°C Xylene 30 min 40°C Xylene 40 min 40°C Wax 70 min 61°C Wax 70 min 61°C Wax 70 min 61°C Deparaffinization of tissue sections
Caution: All steps in this section should be performed in a chemical hood.
Prior to starting the procedure, prepare a series of eleven jars containing the solvent solutions mentioned in each of the steps below.
Place the slides in a slide rack and immerse into a jar of xylene for 5 min.
Immerse in a 2nd jar of xylene for 5 min.
Immerse in a 3rd jar of xylene for 5 min.
Immerse 10 times (2-3 s each time) in a jar of 99.9% ethanol.
Immerse 10 times (2-3 s each time) in a 2nd jar of 99.9% ethanol.
Immerse in a 3rd jar of 99.9% ethanol for 5 min.
Immerse 10 times (2-3 s each time) in a jar of 96% ethanol.
Immerse in a 2nd jar of 96% ethanol for 5 min.
Immerse 10 times (2-3 s each time) in a jar of 70% ethanol.
Immerse in a 2nd jar of 70% ethanol for 5 min.
Immerse in a jar of PBS for 5 min.
Proteinase K treatment
The Proteinase K concentration and length of treatment presented below were optimized for PFA-fixed, paraffin-embedded human embryonic kidney tissue and kidney organoids. For different fixation methods and/or tissue types, these parameters need to be re-optimized (see Note 2).
Add 0.75 μl Proteinase K stock solution per ml of Proteinase K buffer (see Recipe 5) to obtain 1× Proteinase K reagent. For our fixation/tissue combination (Bantounas et al., 2021), we determined the optimum concentration to be 0.33×; therefore, further dilute the 1× Proteinase reagent 1 in 3, with Proteinase K buffer (see Note 2). Make enough working solution for approximately 300 μl of final reagent per section to be stained (although less may be enough in the case of the organoids).
With the slides on a flat surface, apply 300 μl (or enough to cover the section) of Proteinase K reagent directly onto each section.
Transfer the slides to a pre-heated hybridization oven and incubate at 37°C for 10 min (see Note 2).
Place the slides onto a slide rack and wash twice by immersing into a jar of PBS.
Probe hybridization
The final probe concentrations and hybridizing temperature given below are specific for the miRNAs we studied in Bantounas et al. (2021) and are given as examples. These parameters should be optimized separately for each individual miRNA (see Note 3). We recommend that alongside the miRNA of interest, a negative control (scrambled) probe, as well as a positive (U6 snRNA) probe is used (see also Table 2).
Dilute the 2× Formamide-free miRNA ISH buffer with an equal volume of RNase-free water to obtain 1× miRNA ISH buffer. Prepare enough for all probe/section combinations (see Step E2).
In an RNase-free Eppendorf tube, dilute the probe stock in 1× miRNA ISH buffer appropriately to obtain the desired working/final concentration LNA® probe mix (see Note 3). Table 2 shows examples of probes optimized at different final concentrations as used in Bantounas et al. (2021). The final volume should be enough to add 50 μl per section (although you can scale up or down depending on the size of your section).
Table 2. Examples of stock and final concentrations of LNA® probes used on kidney tissue/organoids
Place the tubes in a heating block, at 90°C for 4 min, to denature the probes. Then, centrifuge briefly to collect all the liquid at the bottom of the tube.
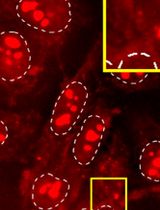
Prepare a slide holding staining tray by placing wet tissue at the bottom and place the slides on top, as shown in Figure 2A.
Apply 50 μl of LNA® probe mix onto each section and replace the lid on the staining tray (Figure 2B).

Figure 2. Set up of slides on staining tray, with wet tissue underneath to prevent evaporation of the applied solution during hybridizationPlace the slide box in the oven at 55°C (see Note 3) for 1 h.
Remove the slide box from the oven, place the slides in a slide rack and immerse in a jar containing 5× SSC buffer (Recipe 6).
Prepare a series of six jars containing different concentrations of SSC buffer (see Recipe 6), as detailed below in Table 3, and wash the slides by serially immersing in each of them for 5 min. Keep the jars in a waterbath set at the hybridization temperature, except the last one (#6), which should be at room temperature.
Table 3. Stringent washes following probe hybridization
Transfer the slides into a jar containing PBS.
Probe Stock concentration Final Concentration Dilution Factor U6 snRNA (+ve control) 0.5 μM 0.1 nM 1: 5,000 miR-199a-3p 25 μΜ 40 nM 1: 625 miR-214-3p 25 μΜ 1 nM 1: 25,000 Scrambled (-ve control) 25 μΜ 40 nM 1: 625 Wash # Buffer 1 5× SSC 2 1× SSC 3 1× SSC 4 0.2× SSC 5 0.2× SSC 6 0.2× SSC Antibody-detection and visualization of bound probe
Caution: In the blocking and immunostaining steps below, do not let the tissue sections dry out, as this can lead to a very high background signal and also risks compromising the integrity of the tissue.
Using the KP marker (see Materials), apply a hydrophobic barrier around each section.
Add 50 μl (or enough to cover the section) blocking solution (Recipe 8) per section and incubate at room temperature for 15 min.
During blocking, dilute the anti-digoxigenin (anti-DIG-AP) antibody 1:400 (see Note 4) in antibody dilution buffer (Recipe 9), allowing 50 μl (or enough to cover the section) per section.
Remove the blocking solution by tipping the slides, then apply the antibody to the sections and incubate with the antibody at room temperature for 1 h.
Wash the slides three times by placing them in a slide rack and successively immersing in three jars containing fresh PBS-T 0.1% (Recipe 7) for 3 min each.
To make the AP reaction substrate, dissolve an NBT-BCIP tablet in 10 ml distilled water. Then, add 20 μl Levamisol stock solution (see Recipes) (2 μl per ml of NBT-BCIP solution).
Caution: The substrate solution must be used immediately and be protected from light.
Apply 50 μl (or enough to cover the section) substrate per section and incubate in the dark for 2 h in a 30°C oven (see Note 5).
To stop the reaction, place the slides in a slide rack and immerse in a jar containing KTBT buffer (Recipe 10) for 5 min.
Repeat the above wash a second time.
Wash the slides with distilled water twice, for 1 min each.
Apply 50 μl (or enough to cover the section) Nuclear Fast Red counterstain for 1 min.
Place the slides in a slide rack and into a jar containing tap water. Rinse under running tap water for 10 min.
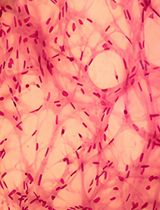
The slides can be examined and imaged using a brightfield microscope. MiRNA-positive cells will be stained blue. Optionally, to capture the entire section in detail, use a 3D-Histech Panoramic-250 microscope slide-scanner. Subsequently, images can be analysed and snapshots captured with the CaseViewer software and further processed using ImageJ. Representative results can be seen in Figure 3.
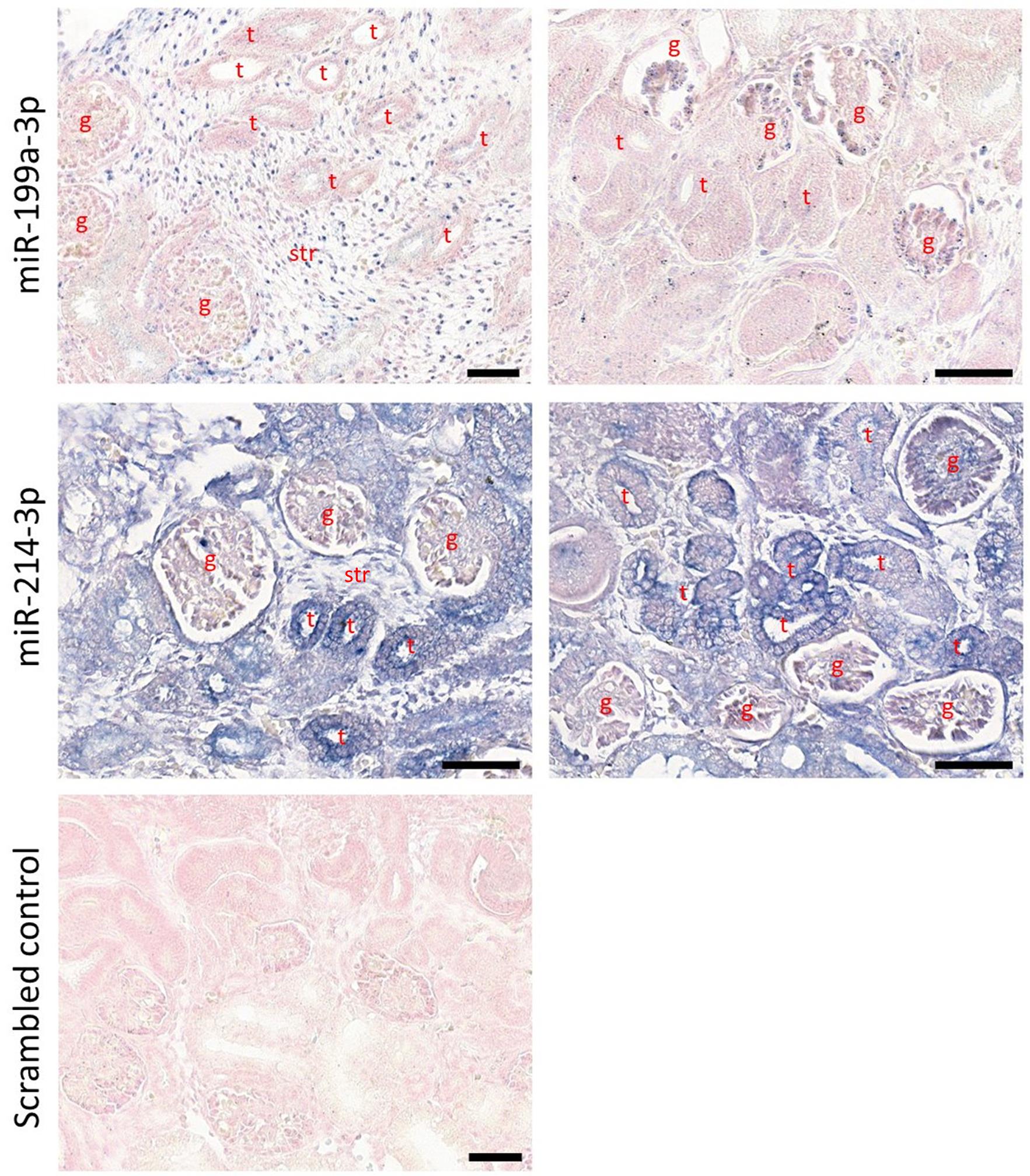
Figure 3. Detection of miRNAs in human fetal kidney sections (week 12 of gestation) by in situ hybridization with digoxigenin-labelled LNA® probes. MiR-199a-3p was present in the stroma and immature glomeruli, while miR-214-3p was present in the stroma, immature glomeruli and tubules (g: glomerulus; t: tubule; str: stroma). Scale bars are 50 μm.
Notes
QIAGEN recommends the use of double-labelled (both 5’ and 3’) digoxigenin probes for miRNA detection. However, we found that in our tissue/miRNA combination, single-labelled (5’) probes gave good results at a lower cost. It is possible that for other tissue/miRNA combinations, a double-labelled probe is indeed necessary (e.g., for less abundant miRNAs).
This protocol assumes PFA fixation and paraffin embedding of tissue sections. If either of these conditions changes, Proteinase K concentration and duration of treatment may need to be re-optimized, even for the same tissue type. In such cases, a pilot experiment must be carried out using the positive control (U6) LNA® probe, varying the range of Proteinase K reagent between 0.03× and 2× and incubating for 10 min. Then, use the concentration that gave the best results and perform a second pilot experiment, varying the duration of treatment between 5 min and 30 min.
The optimal LNA® probe concentration is miRNA-specific, and the optimal hybridization temperature is usually around 30°C below its Tm (as given by the manufacturer). Pilot experiments can be performed for each probe, varying the concentration between 5 nM and 80 nM (although, occasionally, the optimum could be even lower than this; e.g., the miR-214-3p probe in our study was used at 1 nM; Bantounas et al., 2021). The 55°C, mentioned in our protocol, is usually a good starting point as a hybridization temperature, but the optimum can be determined in subsequent pilot experiments, and it usually falls into the 50°C to 60°C range.
Note: It is possible that no positive results are initially obtained in the probe concentration optimization step, in which case, you should optimize the temperature first using a mid-range probe concentration (e.g., 10-20 nM), following which you should refine the concentration further.If desired, the antibody dilution can be optimized by using dilutions in the range of 1:200-1:2,000.
If a 2 h development time results in too strong or too weak a signal, you could vary the incubation time of the AP substrate. With some tissue/miRNA combinations, you may be able to observe the colour as it develops under a light microscope and stop or prolong the incubation time accordingly.
Recipes
4% Paraformaldehyde (PFA) Solution
To a glass beaker, add 800 ml of 1× PBS and heat to 60°C, with stirring on a heated stir plate, in a chemical hood.
Then add 40 g PFA. Raise the pH by adding 10 M NaOH dropwise until the PFA goes into solution (solution becomes clear).
Allow the solution to cool to room temperature and filter (e.g., using a vacuum filter unit).
Adjust the volume to 1,000 ml with 1× PBS.
Check the pH again and adjust with small amounts of 1 M HCl to 6.9.
Store in 10 ml aliquots at -20°C. Defrosted aliquots can be stored at 4°C for up to one month.
1 M Tris Buffer
In 800 ml distilled water, dissolve 121.1 g Tris Base.
Adjust pH to 7.4 by adding concentrated (37% w/v) HCl (approximately 70 ml).
Make sure the solution is at room temperature before making final adjustments to pH, then adjust the total volume to 1,000 ml before autoclaving. Store indefinitely at room temperature.
0.5 M EDTA Solution
To 800 ml of distilled water, add 186.1 g disodium-EDTA2H2O.
While stirring using a magnetic stirrer, adjust the pH to 8, using NaOH. Bring volume to 1,000 ml with water.
Autoclave and store at room temperature. Discard if EDTA precipitates out of solution.
5 M NaCl Solution
Dissolve 292.2 g of NaCl in 800 ml distilled water.
Bring volume to 1,000 ml with water.
Autoclave and store indefinitely at room temperature.
1× Proteinase K Buffer
To 900 ml RNase free water, add:
5 ml of 1 M Tris-HCl, pH 7.4
2 ml 0.5 M EDTA
0.2 ml 5 M NaCl
Bring volume to 1,000 ml, then autoclave. Store indefinitely at room temperature.
SSC Buffer dilutions
5× SSC: Add 250 ml of 20× SSC to 750 ml distilled water
1× SSC: Add 50 ml of 20× SSC to 950 ml distilled water
0.2× SSC: Add 10 ml of 20× SSC to 990 ml distilled water
Autoclave and store indefinitely at room temperature.
PBS-T 0.1%
To 1 L of 1× PBS, add 1 ml Tween-20. Store at room temperature for up to one month.
Blocking solution (prepare fresh each time)
Per 1 ml of PBS-T 0.1% (see Recipe 7 above), add 20 μl sheep serum (2% v/v) and 10 mg BSA (1% w/v). Scale up as necessary.
Antibody dilution buffer (prepare fresh each time)
Prepare PBS-T 0.05% by mixing equal volumes of PBS-T 0.1% (see Recipe 7 above) with PBS.
Then, per 1 ml of the resultant solution, add 10 μl sheep serum (1% v/v) and 10 mg BSA (1% w/v).
KTBT Buffer
To 900 ml RNase free water, add:
7.9 g Tris-HCl
8.7 g NaCl
0.75 g KCl
Bring volume to 1,000 ml with water and autoclave. Store indefinitely at room temperature.
Acknowledgments
The protocol is presented as used in the original research paper (Bantounas et al., 2021), in which the probe-binding and visualizing part is an adaptation, with some modifications, of the protocol recommended in QIAGEN’s miRCURY® LNA® miRNA Detection Probes Handbook (October 2017 edition). The following bodies funded the original research: UK Research and Innovation/Medical Research Council (MRC) UK Regenerative Medicine Platform hub Grant MR/K026739/1; Kidney Research UK John Feehally-Stoneygate Project and Innovation award JFS/RP/008/20160916; Kidneys for Life pump priming grant; Horizon 2020 Marie Skłodowska-Curie Actions Initial Training Network RENALTRACT (642937) grant, EPSRC/MRC Centre for Doctoral Training grant EP/L014904/1. We thank Adrian S. Woolf for providing the human embryonic tissue, which was obtained from the MRC and Wellcome Trust Human Developmental Biology Resource. We also thank the University of Manchester Bioimaging Core Facility for helping us capture the images in Figure 3.
Competing interests
The authors declare no competing interests.
Ethics
Human tissues, collected after maternal consent and ethical approval (REC 08/H0906/21+5), were provided by the MRC and Wellcome Trust Human Developmental Biology Resource (http://www.hdbr.org/).
References
- Bartel, D. P. (2018). Metazoan MicroRNAs. Cell 173(1): 20-51.
- Bantounas, I., Lopes, F. M., Rooney, K. M., Woolf, A. S. and Kimber, S. J. (2021). The miR-199a/214 Cluster Controls Nephrogenesis and Vascularization in a Human Embryonic Stem Cell Model. Stem Cell Reports 16(1): 134-148.
- Bantounas, I., Ranjzad, P., Tengku, F., Silajdzic, E., Forster, D., Asselin, M. C., Lewis, P., Lennon, R., Plagge, A., Wang, Q., Woolf, A. S. and Kimber, S. J. (2018). Generation of Functioning Nephrons by Implanting Human Pluripotent Stem Cell-Derived Kidney Progenitors. Stem Cell Reports 10(3): 766-779.
- Jones, T. F., Bekele, S., O'Dwyer, M. J. and Prowle, J. R. (2018). MicroRNAs in Acute Kidney Injury. Nephron 140(2): 124-128.
- Jørgensen S., Baker A., Møller S., Nielsen B. S. (2010). Robust one-day in situ hybridization protocol for detection of microRNAs in paraffin samples using LNA probes. Methods 52(4): 375-81.
- Lopes, F. M., Roberts N. A., Zeef L. A., Gardiner N. J., Woolf A. S. (2019). Overactivity or blockade of transforming growth factor-β each generate a specific ureter malformation. J Pathol 249(4):472-484.
- Singh S. K., Nielsen P., Koshkin A. A. and Wengel J. (1998). LNA (Locked nucleic acids): synthesis and high-affinity acid recognition. Chem Commun 455-456.
- Takasato, M., Er, P. X., Chiu, H. S., Maier, B., Baillie, G. J., Ferguson, C., Parton, R. G., Wolvetang, E. J., Roost, M. S., Chuva de Sousa Lopes, S. M. and Little, M. H. (2015). Kidney organoids from human iPS cells contain multiple lineages and model human nephrogenesis. Nature 526(7574): 564-568.
- Trionfini, P., Benigni, A. and Remuzzi, G. (2015). MicroRNAs in kidney physiology and disease. Nat Rev Nephrol 11(1): 23-33.
- Ye, J., Bates, N., Soteriou, D., Grady, L., Edmond, C., Ross, A., Kerby, A., Lewis, P. A., Adeniyi, T., Wright, R., Poulton, K. V., Lowe, M., Kimber, S. J. and Brison, D. R. (2017). High quality clinical grade human embryonic stem cell lines derived from fresh discarded embryos. Stem Cell Res Ther 8(1): 128.
- Zhao, H., Ma, S. X., Shang, Y. Q., Zhang, H. Q. and Su, W. (2019). microRNAs in chronic kidney disease. Clin Chim Acta 491: 59-65.
Article Information
Copyright
© 2021 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Lopes, F. M., Kimber, S. J. and Bantounas, I. (2021). In situ Hybridization of miRNAs in Human Embryonic Kidney and Human Pluripotent Stem Cell-derived Kidney Organoids. Bio-protocol 11(17): e4150. DOI: 10.21769/BioProtoc.4150.
Category
Stem Cell > Pluripotent stem cell > Regenerative medicine
Developmental Biology > Cell growth and fate
Molecular Biology > RNA > RNA labeling
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.