- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Liposomal Clodronate-mediated Macrophage Depletion in the Zebrafish Model
(*contributed equally to this work) Published: Vol 11, Iss 6, Mar 20, 2021 DOI: 10.21769/BioProtoc.3951 Views: 6720
Reviewed by: Chiara AmbrogioTianjiao HuangWilliam Jennings Valentine
Original research article
The authors used this protocol in:

Jul 2020

Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Related protocols
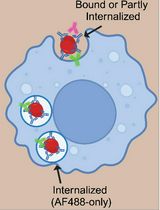

Abstract
The ability to conduct in vivo macrophage-specific depletion remains an effective means to uncover functions of macrophages in a wide range of physiological contexts. Compared to the murine model, zebrafish offer superior imaging capabilities due to their optical transparency starting from a single-cell stage to throughout larval development. These qualities become important for in vivo cell specific depletions so that the elimination of the targeted cells can be tracked and validated in real time through microscopy. Multiple methods to deplete macrophages in zebrafish are available, including genetic (such as an irf8 knockout), chemogenetic (such as the nitroreductase/metronidazole system), and toxin-based depletions (such as using clodronate liposomes). The use of clodronate-containing liposomes to induce macrophage apoptosis after phagocytosing the liposomes is effective in depleting macrophages as well as testing their ability to phagocytose. Here we describe a detailed protocol for the systemic depletion of macrophages in zebrafish larvae by intravenous injection of liposomal clodronate supplemented with fluorescent dextran conjugates. Co-injection with the fluorescent dextran allows tracking of macrophage depletion in real time starting with verifying the successful intravenous injection to macrophage uptake of molecules and their eventual death. To verify a high degree of macrophage depletion, the level of brain macrophage (microglia) elimination can be determined by a rapid neutral red vital dye staining when clodronate injection is performed at early larval stages.
Graphical abstract:
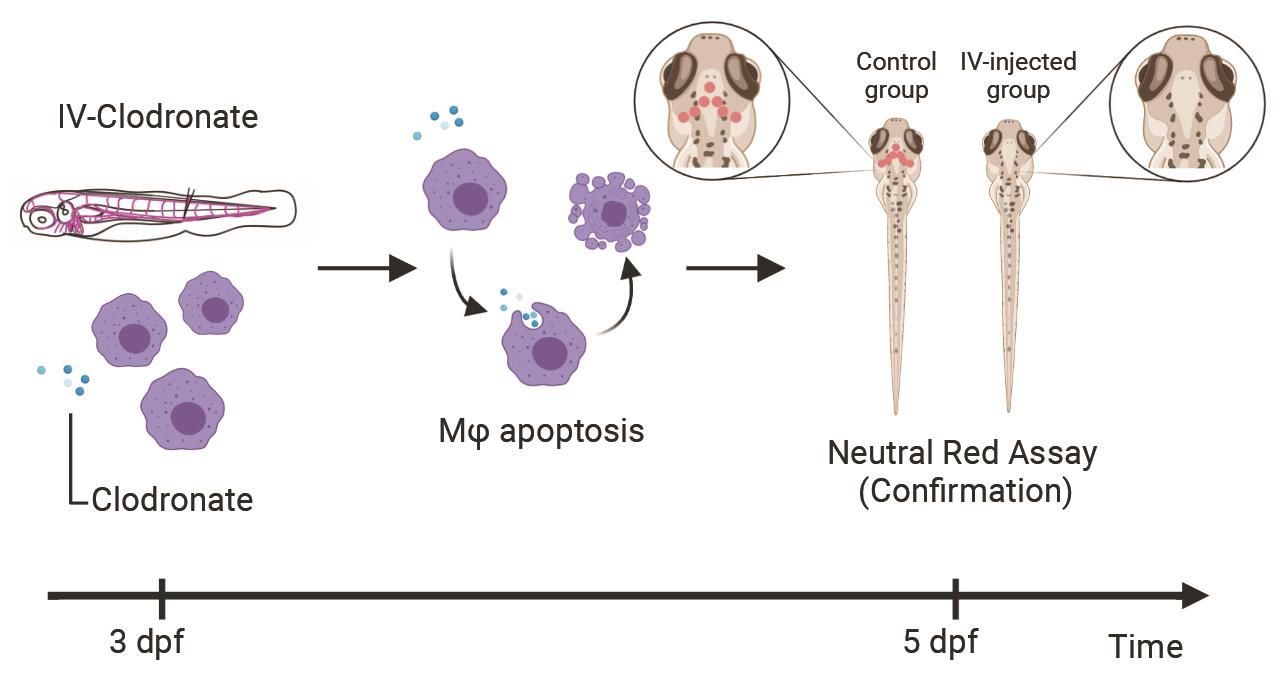
Experimental workflow for in vivo macrophage-specific depletion by liposomal clodronate in larval zebrafish
Background
Macrophages are key constituents of the innate immune system and play important roles in response to infections, sterile inflammation, and environmental changes. One of the most effective ways to uncouple the functions of macrophages from a complex mix of interacting cell types in different physiological contexts is to be able to specifically eliminate macrophages and analyze the phenotypic consequence. Such depletion experiments in mice have provided much insight into the role of macrophages (Hua et al., 2018; Rosowski, 2020). However, our understanding of macrophage functions remains incomplete, and cell depletion experiments in the mouse model are difficult to track and validate in real time. For these reasons, the optical transparency and ease of manipulation of larval zebrafish offer clear advantages for highly traceable and tractable cell ablation in vivo by imaging the target cells in real time and in the whole intact organism. Zebrafish genes and immune system also share a high degree of orthology with those of the human (Yoder et al., 2002; Santoriello et al., 2012; Howe et al., 2013). Furthermore, the adaptive immune system of zebrafish does not become functionally mature until juvenile adult stages (Lam et al., 2004), making the larval zebrafish an excellent platform to study the innate immune system independent of the adaptive immune contributions.
Currently available macrophage depletion methods in zebrafish include genetic and chemogenetic manipulations, and toxin-based depletion. The development of macrophages requires early and continual function of the transcription factor Pu.1 (with the gene name spi1b), together with another transcription factor Irf8 (Li et al., 2011; Shiau et al., 2015; Tenor et al., 2015). Either disruption of pu.1 or irf8 by gene knockout or knockdown by morpholino (MO) anti-sense oligomers, makes a reliable approach for macrophage depletion, while the former ablates myeloid cells, and the latter is more specific to macrophages but also causes an increase in neutrophil numbers (Shiau et al., 2015; Yang et al., 2020). These methods are not amenable to temporal control (Rhodes et al., 2005; Li et al., 2011; Shiau et al., 2015; Rosowski, 2020), while clodronate-mediated depletion of macrophages based on localized microinjection enables some degree of spatial and temporal designation (Bernut et al., 2014).
Clodronate (also known as dichloromethylene diphosphonate) can be metabolized by cells to block mitochondrial respiration due to the formation of a non-hydrolyzable ATP analog, which then causes cell death (apoptosis) (Rosowski, 2020). Once injected as encapsulated in liposomes, clodronate is easily ingested and eliminated by macrophages as it accumulates within the cell (van Rooijen and Hendrikx, 2010). Since neither the clodronate nor the liposomal phospholipids utilized are toxic to other non-phagocytic cells (van Rooijen and Hendrikx, 2010), this approach allows specific depletion of already existing phagocytosing macrophages.
As part of the design for our protocol, we co-injected fluorescently labeled dextran with the liposomal clodronate to allow us to verify a precise and accurate injection and track the effect of the clodronate on macrophages in the entire larvae. To this end, after intravenous co-injection of clodronate liposome with fluorescently tagged dextran, we visually verified the successful injection of the substances into circulation and monitored the uptake of fluorescent dextran by macrophages and their ultimate demise over time. We designed the protocol to include a 48-hour post injection period to allow the effect of clodronate induced apoptosis in macrophages to materialize in light of previous work in chicken and mice showing that the efficacy of clodronate can take a few days depending on the tissue (Kameka et al., 2014; Ponzoni et al., 2018). We confirmed the efficacy of clodronate-mediated macrophage depletion at 48 hours post injection by assessing the remaining number of brain-resident macrophages (microglia), because a rapid analysis of microglia in the living larvae is available by the neutral red vital dye staining. We have chosen to inject at the early larval stage at 3 dpf (days post fertilization) because this is prior to maturation of the blood-brain-barrier (Jeong et al., 2008; O’Brown et al., 2019) at which time we found that our injected substances easily reach macrophages throughout the body including the brain. Using the 48-hour window for clodronate to take effect, we were able to achieve full ablation of microglia in most injected larval zebrafish (Yang et al., 2020). Overall, we found that intravenous microinjection of liposomal clodronate at 3 dpf with a 48-hour incubation time is effective in eliminating macrophages.
Materials and Reagents
1.5 ml microfuge tubes (Eppendorf, SafeLock, catalog number: 00 30120086 )
Polystyrene Petri dish (VWR, catalog number: 25384-342 )
Thin wall borosilicate glass capillaries, 4 inches, OD 1.5 mm with filament (World Precision Instruments, catalog number: TW150F-4 )
Glass bottle
7.5 ml transfer pipettes (VWR, catalog number: 414004-005 )
Low melt agarose (Fisher Scientific, IBI Scientific, catalog number: 50-550-455 ), store at room temperature
PTU (N-Phenylthiourea) (Sigma-Aldrich, catalog number: P7629 ), store at room temperature; made into PTU solution, store at -20 °C
Clodronate Liposomes (Liposoma, catalog number: C41J0781 , https://clodronateliposomes.com), store at 4-7 °C
Control Liposomes (Liposoma, catalog number: B#P37J0718 , https://clodronateliposomes.com), store at 4-7 °C
Dextran, Alexa FluorTM 568: 10,000 MW (Invitrogen, catalog number: D22912 ), store in freezer and protect from light
Neutral Red Dye (Sigma-Aldrich, catalog number: N4638 ), store at room temperature
Tricaine (3-amino benzoic acidethylester) (Sigma-Aldrich, catalog number: A-5040 ) made into tricaine solution, store at -20 °C
50× PTU stock (see Recipes)
25× Tricaine stock solution (100 ml) (see Recipes)
3% Methyl cellulose (see Recipes)
1,000× neutral red solution (see Recipes)
1.5% low melt agarose (see Recipes)
Equipment
Stereomicroscope with LED illuminated base and articulating mirror (World Precision Instruments, model: 504941 , PZMIII-MI)
Fluorescent stereomicroscope fully apochromatic corrected with 16:5:1 zoom optics (Leica, model: M165 MC )
Manual micromanipulator (World Precision Instruments, catalog number: M3301 )
PicoNozzle Kit v2 (World Precision Instruments, catalog number: 5430-ALL )
Pneumatic PicoPump PV 820 (World Precision Instruments, catalog number: SYS-PV820 )
Incubator (Benchmark Scientific, model: H2200-H )
Microelectrode Puller (World Precision Instruments, catalog number: PUL-1000 )
Microwave
Software
Fiji (Schindelin et al., 2012, https://fiji.sc/)
Procedure
Larval zebrafish growth
Incubate zebrafish embryos in fish water in a 28.5 °C incubator.
Conduct a daily water change, and starting at 24 h post-fertilization (hpf), the fish water is also supplemented with PTU at a final concentration of 0.003% to inhibit pigmentation.
PTU (N-Phenylthiourea) inhibits melanogenesis (pigmentation) by blocking all tyrosinase-dependent steps in the melanin pathway (Karlsson et al., 2001).
Check the health status of the zebrafish larvae before injection. By 3 dpf healthy larvae will have a consistent rhythmic heartbeat with apparent blood flow, intermittent bursts of movement, and a straight body.
Prepare liposome mix with fluorescent dextran
Use clodronate liposomes and control liposomes directly at 5 mg/ml at the concentration they were delivered in liquid form from the manufacturer. We recommend making aliquots of the liposomes for storage at 4 °C to eliminate possible contamination from multiple usage; we use the aliquots within 12 months of storage. Supplement the liposome with Alexa 568 labeled dextran at 1:100 dilution of a 5 ng/nl stock for visualization of the injection.
For example: add 0.5 μl of a 1:10 dilution of Alexa 568 labeled dextran at 5 ng/nl to 4.5 μl of clodronate liposomes at its original concentration of 5 mg/ml.
Vortex to mix until a homogenous mixture for about 30-60 s.
Needle and microinjector set-up
Review the manufacturer’s recommendations and instructions for the micro-needle puller and the microinjector before operating.
Use a micropipette needle puller to make a fine microinjection needle (refer to Table 1 and Figure 1).
Microelectrode puller program (see Table 1)
Table 1. 4-step protocol for glass capillary tube pullingStep Heat Force Distance Delay 1 690 260 7.3 0 2 500 240 0.5 4 3 500 230 0.5 10 4 380 240 0.5 20 
Figure 1. Pulling microinjection needle. A. Capillary tube loaded into micro-needle puller. B. Magnified view of heated filament surrounding glass capillary. C. Creation of 2 microinjection needles from pulling (double-sided white arrow).Load 3 μl of injection material into the wide end of the microcapillary glass tube.
Shake the capillary tube to bring the injection material to the tip of the needle and to remove any air bubbles.
Turn on the air source and the microinjector, and transfer the needle into the microinjector.
Break the tip of the microinjection needle using forceps to an extent that allows the injection materials to release consistently and minimize the needle puncture on the target tissue.
Adjust the pressure on the pneumatic pump and the location of microinjection syringe pump to ensure consistent 0.5 nl per foot-pedal kick of injection so each injection consists of two kicks per larval fish for a total of 1 nl.
Size of each injection kick can be estimated by injecting into air which creates a spherical drop at the needle tip. The diameter of the liquid sphere of about 100 μm is estimated using a stage micrometer to give 0.5 nl volume.
Adjust the micromanipulator so that the needle can reach the center of the stage and touch the base so that the needle can reach any mounted larvae in your field of view under microscope.
Mounting zebrafish for microinjections
Use a microwave to melt 1.5% low-melt agarose (for a short time, 10-20 s). See Recipes for low-melt agarose preparation.
Use a plastic transfer pipette to transfer multiple larvae at 3 dpf to the center of a 10 cm Petri dish lid. Many larvae can be transferred at once upwards of 10-15 larvae.
Remove as much fish water as possible to leave only minimal water around fish as to not dehydrate them, then slowly pipette in lukewarm low melt agarose and gently swirl the larvae to mix them into the agarose (or by pipetting them up and down 1-2 times).
The agarose will be very hot immediately after heating. To prevent scalding and killing the larvae, wait until the agarose is not hot to the touch, but still warm and fluid. The low-melt agarose will remain fluid at 37 °C and set rapidly below 25 °C. So you want to aim to use the agarose at the temperature between 30 °C and 40 °C. You can also keep the agarose on a heat block at 60 °C so it remains in solution for a longer period of time as you work on mounting the larvae. You can expedite the cooling process by pipetting hot agarose up and down on a clean surface (such as in another dish).
The goal is to have the larvae embedded in a thin layer of agarose so that when it comes to using the fine glass needle for injection, the needle does not need to penetrate through much gel to reach your desired tissue target (see Figure 2). Too much agarose will make it hard to direct the needle to your target site without having it bend or swerve in the agarose.

Figure 2. Set-up of zebrafish larval microinjections. A. Mounted larvae are placed under a fluorescent stereomicroscope adjacent to a microinjection needle attached to a micro-manipulator. B. Higher magnification of mounted larvae. Dotted circle shows solidified thin layer of low-melt agarose covered with a small pool of system water supplemented with PTU. Black arrows point to larvae. Red arrow points to tip of needle (Note: This arrow is not pointing to the injection site.)The most time-sensitive step is in orienting all the zebrafish larvae quickly in the agarose before it begins to re-solidify. You can expect to have less than 1-2 min for this step. Use fine forceps to quickly orient each one on their sides, but without concern of the exact orientation or body alignment, only that the tail region containing the caudal vein plexus is accessible to your needle. Take caution to not poke the larvae to cause injury but to use the agarose around the larvae to move them.
Wait for agarose around the larvae to cool down and solidify before starting injections.
Add a small amount of fish system water to cover the agarose to prevent dehydration and keep the larvae healthy during the operation (see Figure 2).
Intravenous microinjections
Use the foot pedal to inject 1 nl of clodronate liposomes or control liposomes into caudal vein plexus, which is located in the tail tissue right past the yolk extension (see Graphical abstract).
Validate each injection by seeing an immediate distribution of the injected fluorescent dextran mix into the heart and subsequent circulation throughout the body vasculature using a fluorescent stereomicroscope. Any larvae after injection that do not show the correct distribution of the fluorescent marker in the body vasculature is removed from the dish using forceps.
Gently recover the injected fish from the agarose into fresh fish water supplemented with 0.003% PTU and transfer them back into the 28.5 °C incubator.
Use the side of the forceps to create a break in the agarose beginning underneath the head and dragging the forceps along the body until the tip of the tail is reached. Larvae should be able to squirm out easily and swim into the system water surrounding the agarose. Pipette released larval zebrafish into the dish with fresh fish water.
After microinjection, monitor all the fish for normal health and behavior. Remove any sickly or abnormal fish, and only continue processing the healthy and normal larvae.
Neutral red staining
In a Petri dish, stain a subset of your injected larvae at 48 hpi (hours post injection) with neutral red at 2.5 μg/ml in fish water supplemented with 0.003% PTU at room temperature for 45 min to 1 h (see Figure 3A).
Remove the majority of the neutral red supplemented fish water by exchanging it for fresh new fish water supplemented with PTU (see Figure 3B).
Leave the larvae in incubator for another 2-3 h before analyzing under a stereomicroscope.
Before analyzing, anesthetize larvae with tricaine at 0.5× strength, which the larvae can stay in for up to 45 min with the ability to fully recover back to normal and moving again. For the anesthetics to be working, larvae should not be moving so you can use them for your analysis.
Note: Please follow your institutional guidelines on the appropriate protocol for anesthetizing zebrafish.
Mount larvae in 3% methyl cellulose to position the larvae for analyzing and imaging under a stereomicroscope with a color camera. The larvae should be mounted brain side up to image the microglia under an upright stereomicroscope (see Figure 3C-3E).
Notes:
You can mount larvae on a dish or any flat platform to visualize them under a stereoscope.
Because methyl cellulose is highly viscous and these larvae were temporarily anesthetized, they remain sufficiently still for quick imaging without further anesthesia in only methyl cellulose for at least up to 10 min.
Depending on the endpoint of the experiment, these larvae can be directly processed after imaging for genotyping, or recovered by washing them 1-2 times in clean fish water to remove the methyl cellulose at which point they can return to their growing chamber (an incubator) or fixed for further analysis. Fish larvae should remain well and alive during this entire period up to the endpoint processing.
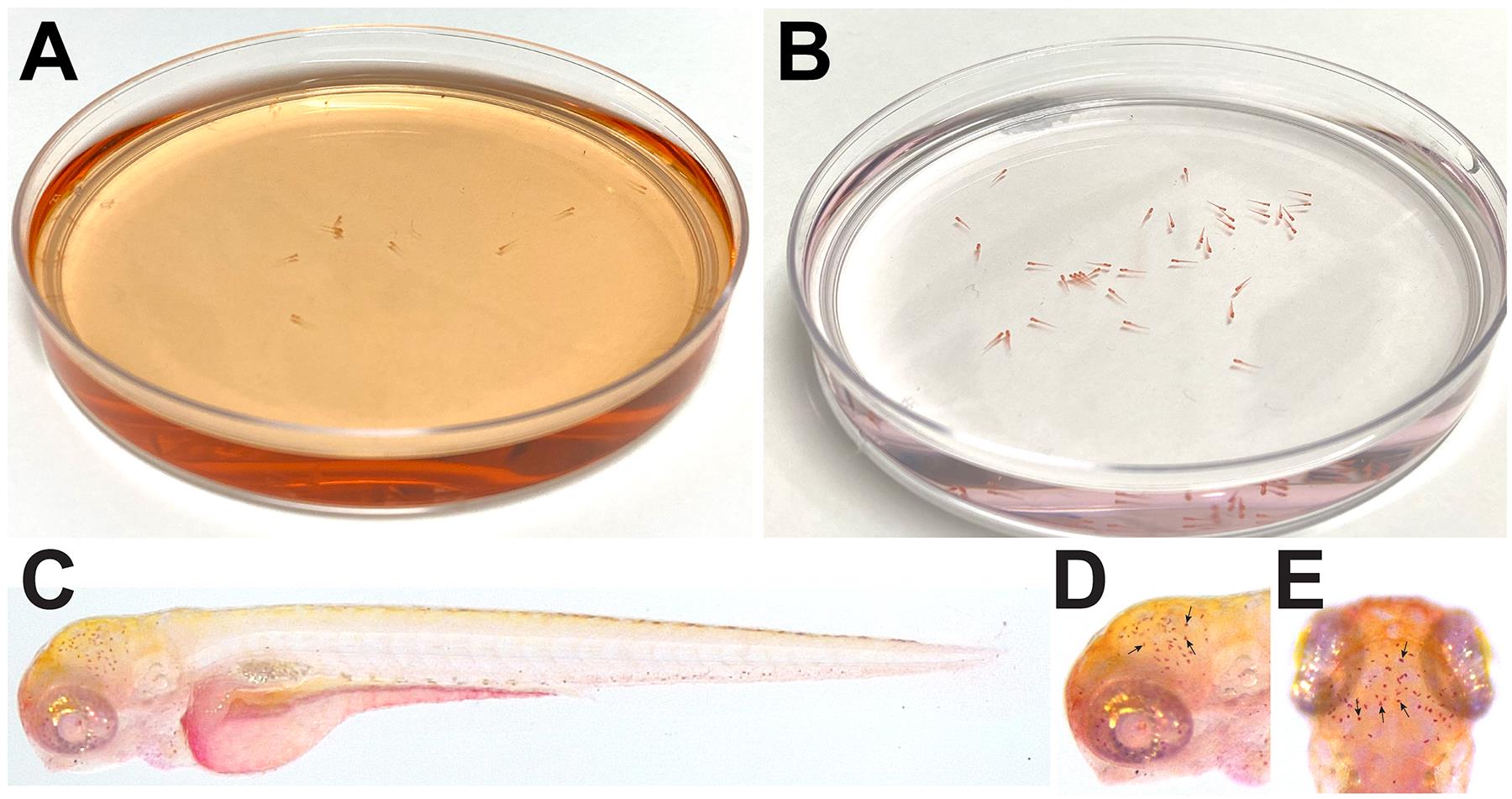
Figure 3. Neutral red analysis. A. Color of fish water after addition of neutral red. B. Color of fish and water after removal of neutral red. C. Full body image of control uninjected 4 dpf larvae after neutral red staining. D and E. Lateral (D) and dorsal (E) view of neutral red staining. C-E. Larvae imaged in 3% methyl cellulose. Black arrows point to individual microglial cells.
Data analysis
The intravenous administration of clodronate liposomes offer the ability to deplete macrophages throughout the body, including microglia and peripheral macrophages, without the maintenance of a transgenic line. Analyze the efficiency of clodronate-mediated macrophage depletion by comparing the number of microglia in the brains of liposomal clodronate injected group with the control liposomal vehicle or control uninjected group using the neutral red vital dye assay at 48 hpi (see Figure 4–Figure Supplement 2 in Yang et al., 2020).
Recipes
50× PTU stock (1 L)
Dissolve 1.5 g N-Phenylthiourea (PTU) in 1 L of distilled water by stirring overnight at room temperature.
Aliquot and store concentrated stock at -20 °C.
To inhibit pigmentation in developing embryos,use at a 1× concentration in fish water and replace daily.
1× is 0.003% [weight (g) by volume (per 100 ml)] in fish water.
25× Tricaine stock solution (100 ml)
Add 400 mg of Tricaine powder (3-amino benzoic acid ethyl ester) to 100 ml of distilled water. Store concentrated stock at -20 °C. To use tricaine as an anesthetic,use at a 0.5-1.0× concentration.
3% Methyl cellulose
Note: You will need 3 g of methyl cellulose powder in 100 ml of distilled water.
To dissolve methyl cellulose powder in water, bring half of the distilled water to 60 °C. The remaining volume of water should be stored at 4 °C and left to cool for 30 min.
Add 3 g of methyl cellulose powder to boiling water and stir. The mixture will become extremely viscous quickly.
Remove the solution from the heat as soon as stirring becomes difficult.
Add the chilled water and continue to stir for 30 s.
To fully dissolve particles and remove bubbles the solution must be placed at -20 °C overnight. To prevent glass from breaking, gradually lower the temperature by placing glass at 4 °C for 1-2 h before placing at -20 °C overnight.
1,000× neutral red dye solution
Dissolve neutral red powder in distilled water at a concentration of 2.5 mg/ml for a 1,000× stock. For live staining of microglia in zebrafish, use neutral red at 1× strength.
1.5% low melt agarose
Dissolve 1.5 g of low melt agarose in 100 ml distilled water using the microwave for less than 30 s.
Monitor the heating process as low melt agarose can easily overheat and bubble over.
Once this solution is made, it can be stored in a glass bottle with a lid at room temperature and be repeatedly re-used by re-melting the agarose in the microwave.
Acknowledgments
The graphical abstract was created with BioRender.com. This protocol accompanies the publication (Yang et al., 2020). The work was funded by NIH NIGMS grant 1R35GM124719 to C.E.S.
Competing interests
The authors declare no competing interests.
Ethics
Animal experimentation: This study was performed in strict accordance with the approved institutional animal care and use committee (IACUC) protocols (#16-160 and #19-132) of the UNC Chapel Hill.
References
- Bernut, A., Herrmann, J. L., Kissa, K., Dubremetz, J. F., Gaillard, J. L., Lutfalla, G. and Kremer, L. (2014). Mycobacterium abscessus cording prevents phagocytosis and promotes abscess formation. Proc Natl Acad Sci U S A 111(10): E943-952.
- Howe, K., Clark, M. D., Torroja, C. F., Torrance, J., Berthelot, C., Muffato, M., Collins, J. E., Humphray, S., McLaren, K. and Matthews, L. et al. (2013). The zebrafish reference genome sequence and its relationship to the human genome. Nature 496(7446): 498-503.
- Hua, L., Shi, J., Shultz, L. D. and Ren, G. (2018). Genetic models of macrophage depletion. Methods Mol Biol 1784: 243-258.
- Jeong, J. Y., Kwon, H. B., Ahn, J. C., Kang, D., Kwon, S. H., Park, J. A. and Kim, K. W. (2008). Functional and developmental analysis of the blood-brain barrier in zebrafish. Brain Res Bull 75(5): 619-628.
- Kameka, A. M., Haddadi, S., Jamaldeen, F. J., Moinul, P., He, X. T., Nawazdeen, F. H., Bonfield, S., Sharif, S., van Rooijen, N. and Abdul-Careem, M. F. (2014). Clodronate treatment significantly depletes macrophages in chickens. Can J Vet Res 78(4): 274-282.
- Karlsson, J., von Hofsten, J. and Olsson, P. E. (2001). Generating transparent zebrafish: a refined method to improve detection of gene expression during embryonic development. Mar Biotechnol (NY) 3(6): 522-527.
- Lam, S. H., Chua, H. L., Gong, Z., Lam, T. J. and Sin, Y. M. (2004). Development and maturation of the immune system in zebrafish, Danio rerio: a gene expression profiling, in situ hybridization and immunological study. Dev Comp Immunol 28(1): 9-28.
- Li, L., Jin, H., Xu, J., Shi, Y. and Wen, Z. (2011). Irf8 regulates macrophage versus neutrophil fate during zebrafish primitive myelopoiesis. Blood 117(4): 1359-1369.
- O'Brown, N. M., Megason, S. G. and Gu, C. (2019). Suppression of transcytosis regulates zebrafish blood-brain barrier function. Elife 8: e47326.
- Ponzoni, M., Pastorino, F., Di Paolo, D., Perri, P. and Brignole, C. (2018). Targeting macrophages as a potential therapeutic intervention: impact on inflammatory diseases and cancer. Int J Mol Sci 19(7).
- Rhodes, J., Hagen, A., Hsu, K., Deng, M., Liu, T. X., Look, A. T. and Kanki, J. P. (2005). Interplay of pu.1 and gata1 determines myelo-erythroid progenitor cell fate in zebrafish. Dev Cell 8(1): 97-108.
- Rosowski, E. E. (2020). Determining macrophage versus neutrophil contributions to innate immunity using larval zebrafish. Dis Model Mech 13(1): dmm041889.
- Santoriello, C. and Zon, L. I. (2012). Hooked! Modeling human disease in zebrafish. J Clin Invest 122(7): 2337-2343.
- Schindelin, J., Arganda-Carreras, I., Frise, E., Kaynig, V., Longair, M., Pietzsch, T., Preibisch, S., Rueden, C., Saalfeld, S., Schmid, B., Tinevez, J. Y., White, D. J., Hartenstein, V., Eliceiri, K., Tomancak, P. and Cardona, A. (2012). Fiji: an open-source platform for biological-image analysis. Nat Methods 9(7): 676-682.
- Shiau, C. E., Kaufman, Z., Meireles, A. M. and Talbot, W. S. (2015). Differential requirement for irf8 in formation of embryonic and adult macrophages in zebrafish. PLoS One 10(1): e0117513.
- Tenor, J. L., Oehlers, S. H., Yang, J. L., Tobin, D. M. and Perfect, J. R. (2015). Live imaging of host-parasite interactions in a zebrafish infection model reveals cryptococcal determinants of virulence and central nervous system invasion. mBio 6(5): e01425-01415.
- van Rooijen, N. and Hendrikx, E. (2010). Liposomes for specific depletion of macrophages from organs and tissues. Methods Mol Biol 605: 189-203.
- Yang, L., Jimenez, J. A., Earley, A. M., Hamlin, V., Kwon, V., Dixon, C. T. and Shiau, C. E. (2020). Drainage of inflammatory macromolecules from the brain to periphery targets the liver for macrophage infiltration. Elife 9: e58191.
- Yoder, J. A., Nielsen, M. E., Amemiya, C. T. and Litman, G. W. (2002). Zebrafish as an immunological model system. Microbes Infect 4(14): 1469-1478.
Article Information
Copyright
![]() Yang et al. This article is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0).
Yang et al. This article is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0).
How to cite
Readers should cite both the Bio-protocol article and the original research article where this protocol was used:
- Yang, L., Rojas, A. M. and Shiau, C. E. (2021). Liposomal Clodronate-mediated Macrophage Depletion in the Zebrafish Model. Bio-protocol 11(6): e3951. DOI: 10.21769/BioProtoc.3951.
- Yang, L., Jimenez, J. A., Earley, A. M., Hamlin, V., Kwon, V., Dixon, C. T. and Shiau, C. E. (2020). Drainage of inflammatory macromolecules from the brain to periphery targets the liver for macrophage infiltration. Elife 9: e58191.
Category
Immunology > Immune cell function > Macrophage
Cell Biology > Cell-based analysis
Biological Sciences > Biological techniques
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.