- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Permethylation and Microfractionation of Sulfated Glycans for MS Analysis

Published: Vol 10, Iss 10, May 20, 2020 DOI: 10.21769/BioProtoc.3617 Views: 4819
Reviewed by: Manjula MummadisettiAnonymous reviewer(s)
Original research article
The authors used this protocol in:

Sep 2018
Advertisement



Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics







Abstract
Sulfated glycans are barely detectable in routine mass spectrometry (MS)-based glycomic analysis due to ion suppression by the significantly more abundant neutral glycans in the positive ion mode, and sialylated non-sulfated glycans in the negative ion mode, respectively. Nevertheless, the negative charge imparted by sulfate can be advantageous for selective detection in the negative ion mode if the sialic acids can first be neutralized. This is most conveniently achieved by a concerted sample preparation workflow in which permethylation is followed by solid phase fractionation to isolate the sulfated glycans prior to MS analysis. Importantly, we demonstrated that conventional NaOH/DMSO slurry permethylation method can retain the sulfates. Instead of extracting permethylated glycans into chloroform for sample clean-up, reverse phase C18 cartridge coupled with self-packed amine-tip or mixed mode weak anion exchange cartridge can be utilized to obtain in good yield the non-sulfated, mono-sulfated, and multiply sulfated permethylated glycans in separate fractions for sulfoglycomic analysis.
Keywords: Permethylated sulfated glycansBackground
Sulfated glycans carrying sulfo sialyl LeX glycotope are ligands of L-selectin involved in lymphocyte homing in physiological and inflammatory states (Rosen, 2004; Kawashima, 2006). Many glycan array studies also showed that sulfated glycotopes are better ligands of several galectins (Ideo et al., 2002) and Siglecs (Bochner et al., 2005). However, there is very limited knowledge of endogenous sulfated glycans expressed on different types of mammalian cells, tissues or organism due to their low abundance and availability of few well-defined antibodies. Advanced mass spectrometry (MS)-based analysis is the only viable technique that would afford the requisite high sensitivity and accuracy for a meaningful probing of the cellular sulfoglycome.
Sulfated glycans are not readily detected in the conventional MS-based glycomic profiling. For analysis of non-derivatized, native sulfated glycans in the negative ion mode, the main problem is that sialylated glycans also carry negative charges and these are usually more abundant. Removing the sialic acids by treatment with sialidase is commonly applied (Hernandez Mir et al., 2009), but information on the sulfated sialylated glycans, which maybe the true physiological ligands for endogenous lectins is lost. For permethylated glycans, the advantages are neutralization of the negative charge of sialic acid and more reliable, sequence informative fragment ions can be obtained by MS/MS analyses. A precondition though is that the widely used sample clean-up method following permethylation, such as extraction by chloroform/water partition, which would result in loss of permethylated sulfated glycans into the aqueous layer, should be avoided (Morelle and Michalski, 2007; Kumagai et al., 2013).
We have shown that the conventional NaOH/DMSO slurry permethylation can retain the sulfates, and reverse phase (C18) solid phase extraction cartridge can be used to replace chloroform extraction (Yu et al., 2009; Khoo and Yu, 2010). Both permethylated non-sulfated and sulfated glycans are retained, extensively washed, and then eluted without introducing salts. MALDI-MS screening of permethylated glycans in the negative ion mode gives a quick survey for the presence of sulfated glycans. Instead of using 2,5 dihydroxybenzoic acid (DHB), which is a common matrix for glycan analysis in the positive ion mode (Harvey, 1993), we found that 3,4-diaminobenzophenone (DABP) afforded a better sensitivity for detecting permethylated sulfated glycans in the negative ion mode. Non-sulfated, mono-sulfated, di-sulfated permethylated glycans can be further fractionated by using a self-packed amine tip. Alternatively, a mixed mode weak anion exchange (MAX) cartridge containing both reverse phase and weak anion exchange properties can be used to replace the two-step process of cleaning and fractionation (Cheng et al., 2013; Cheng et al., 2015; Hsiao et al., 2017). The developed workflow for sulfoglycomics (Figure 1) has been successfully applied to many different biological samples such as lymph nodes from different glycosyltransferase/sulfotransferase knockout mice (Mitoma et al., 2007; Patnode et al., 2013), ovarian cancer (Shibata et al., 2012; Yu et al., 2013), colon cancer (Yu et al., 2018), immune cells (Patnode et al., 2013; Wang et al., 2013). Collectively, it showed that sulfated N- and O-glycans have a wider and ubiquitous occurrence than previously appreciated.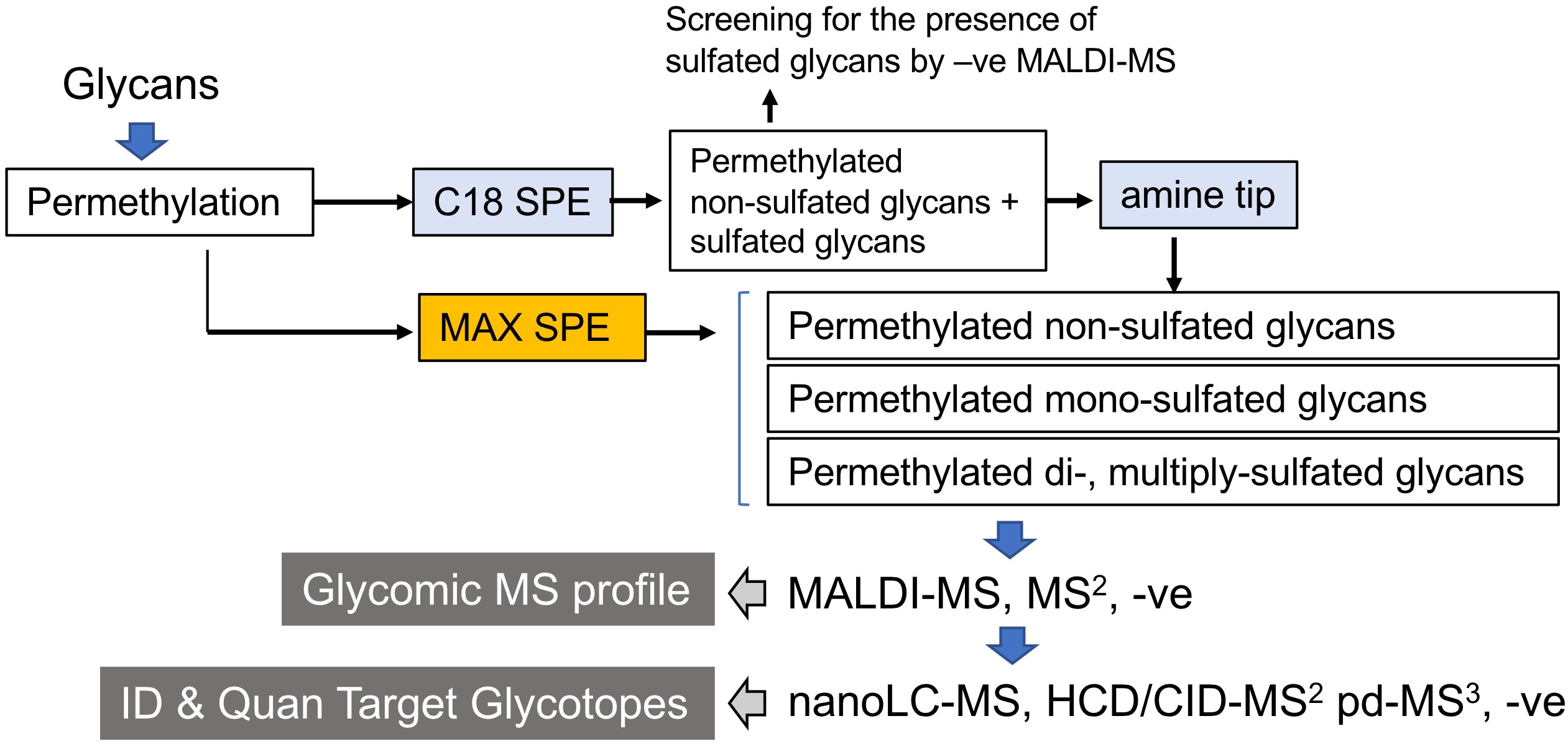
Figure 1. Schematic workflow for MS-based sulfoglycomic analyses. The key steps are reverse phase C18 which retains all permethylated glycans, and further fractionation of sulfated permethylated glycans from non-sulfated ones by amine beads. Alternatively, a mixed mode anion exchange (MAX) SPE is equally effective. MALDI-MS screening in the negative ion mode affords a quick profile of the sulfoglycome. This protocol is applicable to all commonly found sulfated N-glycans and O-glycans, irrespective of their reducing end status. The procedure for additional nanoLC-nanoESI-MS/MS analyses was described in another paper entitled “Negative Ion Mode nanoLC-ESI-MS/MS Analyses of Permethylated Sulfated Glycans” in Bio-protocol (Yu et al., 2020). SPE, solid phase extraction.
Materials and Reagents
- C18 cartridge (Waters, Sep-Pak, catalog number: WAT091139 )
- ZipTipC18 (Merck Millipore, catalog number: ZTC18S096 )
- NUCLEOSIL® 100-5 NH2 (MACHEREY-NAGEL, Nucleosil, catalog number: 712200.1 0)
- Disposable 20 ml Syringe (BD, catalog number: 300296 )
- Filter paper
- Oasis MAX 3 cc Vac Cartridge (Waters, Oasis, catalog number: 186000367 )
- Several clear chemical glass bottles, 250 ml (PYREX®, catalog number: 1515-06D ), 500 ml (PYREX®, catalog number: 1515-08D )
- PYREX® 13 x 100 mm round bottom glass tubes (Corning, catalog number: 99449-13 )
- Corning® Reusable Phenolic GPI 13-415 screw cap with PTFE Liner (Corning, catalog number: 9998-13 )
- Glass Pasteur pipette (Corning, catalog number: 7095B-9 ) and plastic pipette tips
- pH-indicator strips pH 0-14 Universal indicator (Merck Millipore, catalog number: 1.09535.0001 )
- 1.5 ml Eppendorf safe-lock tubes (Eppendorf, catalog number: 00 30120086 )
- Sodium hydroxide (NaOH) (Merck Millipore, catalog number: 106498 )
- Dimethyl sulfoxide (DMSO) (Merck Millipore, catalog number: 1029310161 )
- Methyl iodide (stabilized with silver) (Merck Millipore, catalog number: 806064 )
- 100% ethanoic acid (glacial) (Merck Millipore)
- Acetonitrile (ACN) (VWR chemicals, catalog number: 20060.32 0)
- Methanol (VWR chemicals, catalog number: BDH1135-1LP )
- 3,4-diaminobenzophenone (DABP) (Acros OrganicsTM, catalog number: 184800250 )
- Ammonium acetate (NH4OAc) (Sigma Aldrich, catalog number: 5.43834 )
- Trifluoroacetic acid (TFA) (Sigma-Aldrich, catalog number: 302031-100m ), store in 4 °C
- Formic acid (FA) (Merck Millipore, catalog number: 5.438040.100 )
- 2.5% ACN (200 ml) (see Recipes)
- 10% ACN (200 ml) (see Recipes)
- 25% ACN (200 ml) (see Recipes)
- 50% ACN (200 ml) (see Recipes)
- 0.1% TFA (50 ml) (see Recipes)
- 75% ACN, 0.1% TFA (50 ml) (see Recipes)
- 95% ACN (50 ml) (see Recipes)
- 95% ACN, 0.1% FA (50 ml) (see Recipes)
- 50% ACN, 0.1% FA (50 ml) (see Recipes)
- 1 M ammonium acetate (NH4OAc) (see Recipes)
- 50% ACN, 2.5 mM NH4OAc (50 ml) (see Recipes)
- 50% ACN, 10 mM NH4OAc (50 ml) (see Recipes)
- 100 mM NH4OAc (200 ml) (see Recipes)
- 80% ACN, 1 mM NH4OAc (200 ml) (see Recipes)
- 60% ACN, 20% methanol, 100 mM NH4OAc (200 ml) (see Recipes)
Equipment
- Ceramic mortar and pestle
- Laboratory fume hood
- 50 °C heater
- Vortex-Genie® 2 (Scientific Industries) coupled with 9-16 mm foam inserts (Scientific Industries, model: 503-0278-00 )
- Savant SpeedVac® concentrator (Thermo Scientific, model: ISS110 ) with rotor (Thermo Scientific, catalog number: RH20-12 )
- MALDI-TOFTOF 4700 Proteomics Analyzer or any equivalent MALDI-TOFTOF series (Applied Biosystems, Sciex) mass spectrometer
Software
- Data Explorer version 4.5 (Applied Biosystems)
Procedure
- Permethylation of glycans
- Any glycan of reasonable size including the normal range of mammalian N- and O-glycans prepared from glycoprotein(s), cells, tissues and organs using the conventional protocols (Jang-Lee et al., 2006; Morelle and Michalski, 2007), with free or reduced reducing ends depending on the glycan release methods, can be subjected to the permethylation and sample clean-up/ microfractionation protocols described here. Heavily sulfated glycosaminoglycan fragments have not been tested but 50 pmole starting material of tetrasulfated diLacNAc tetrasaccharides
could be efficiently permethylated, retained by SepPakC18, and successfully detected by subsequent MALDI-MS analysis (Yu et al., 2009). - Aliquot the suitably prepared glycan sample (in Milli-Q water) into a 13 x 100 mm glass tube.
- Dry the glycans by SpeedVac.
- Add 5 ml of DMSO into mortar followed by 5 NaOH pellets. Grind the NaOH pellets to a slurry by the use of a pestle.
- Add 0.2 ml of the NaOH/DMSO slurry into glass tube containing glycans, followed by the addition of 0.1 ml of methyl iodide. Perform this step in the laboratory fume hood.
- Put the sample onto vortex mixer and vortex gently for 3 h, at 4 °C
- Add dropwise 0.2 ml of cold Milli-Q water into glass tube directly to quench the reaction, followed by careful neutralization with several drops of 5% aqueous acetic acid. Check the pH value of sample until pH 5-8 by pH indicator strips. Remove methyl iodide by putting samples under a stream of nitrogen in the fume hood.
Note: The neutralized reaction mix can be loaded into Oasis® MAX cartridge directly (see Procedure E).
- Any glycan of reasonable size including the normal range of mammalian N- and O-glycans prepared from glycoprotein(s), cells, tissues and organs using the conventional protocols (Jang-Lee et al., 2006; Morelle and Michalski, 2007), with free or reduced reducing ends depending on the glycan release methods, can be subjected to the permethylation and sample clean-up/ microfractionation protocols described here. Heavily sulfated glycosaminoglycan fragments have not been tested but 50 pmole starting material of tetrasulfated diLacNAc tetrasaccharides
- Clean-up permethylated glycans by Sep-Pak C18
- Equilibrate C18 Sep-Pak cartridge by 3 ml of 100% ACN, 3 ml of methanol, 3 ml of water sequentially.
- Transfer the neutralized reaction mixture into the primed C18 Sep-Pak cartridge.
- Wash cartridge with 5 ml of water, 5 ml of 2.5% ACN, and 10% ACN subsequently to remove hydrophilic salts and contaminants.
- Elute permethylated glycans by 3 ml of 25% ACN, followed by 3 ml of 50% ACN into the same tube.
Notes:- Di-sulfated permethylated N-glycans will be collected in the 25% ACN fraction, and mono-sulfated along with non-sulfated permethylated N-glycans will be eluted in 50% ACN fraction.
- For O-glycans, di-sulfated, mono-sulfated, part of smaller non-sulfated permethylated O-glycans will normally be found in 25% ACN fraction, whereas larger mono-sulfated permethylated O-glycans and larger non-sulfated O-glycans will be collected in 50% ACN fraction.
- Dry samples by SpeedVac
- Screen permethylated sulfated glycans by MALDI-MS in negative ion mode
- Prepare 10 mg/ml of 3,4-diaminobenzophenone in 75% ACN, 0.1% TFA in a 1.5 ml Eppendorf tube.
- Dissolve the permethylated glycan samples in 20-50 μl of ACN.
Note: The volume of ACN to dissolve the sample highly depends on the amount of glycans. If the quantity is unclear, 20-50 μl of ACN is a good starting volume. - Take 0.5 μl of sample and mix well with 0.5 μl of 3,4-DABP as matrix in an Eppendorf tube.
- Spot the mixed sample onto the MALDI target plate above a 50 °C heater.
- Acquire MS data in the negative ion mode, reflector mode.
- MS data was acquired under either manual or automated target movement depending on the state of crystallization
- The laser energy set for 3,4-DABP is similar to that for 2,5-DHB. In our 4700 Proteomics Analyzer and experience on 4800 Plus MALDI TOF/TOF Analyzer, the laser energy is set to around 4,500-5,000. Laser Nd-YAG 200 Hz is used.
- A typical acquisition comprises a total of 20 sub-spectra with 50 laser shots per sub-spectrum.
- Separation of sulfated permethylated glycans from non-sulfated ones by amine tip
- Prepare self-packed amine tip.
- Take a 200 μl yellow pipette tip and plug its tapered end with filter paper (0.1 cm x 0.1 cm size) by the use of 5 cm length needle or any equivalent tool (as shown in Figure 2A).
- Take up the amine beads in methanol, and pack them into the yellow tip. The volume of the packed beads in yellow pipette tip is normally 20 μl.
- Use the plunger of a 20 ml syringe (Figure 2B) to gently apply pressure to wash out the methanol (Figure 2C). Do not completely dry out the self-packed amine tip.
- Equilibrate the packed amine tip by adding 100 μl of 100% ACN and again use the 20 ml syringe plunger to apply pressure to push out the solvent. Follow with another 100 μl of 95% ACN with 0.1% FA and 100 μl of 50% ACN with 0.1% FA, and 100 μl of 95% ACN with 0.1% FA, sequentially.
- Dissolve the permethylated glycans from Step B5 in 20-50 μl of 100% ACN.
- Load the permethylated glycans onto self-packed amine tip and collect the flow through in the Eppendorf tube.
- Reload the collected flow through sample onto tip, and collect the flow through in the new Eppendorf tube.
- Wash the tip with 100 μl of 95% ACN. Collect the flow through and wash fractions in the same Eppendorf tube. This tube contains non-sulfated permethylated glycans.
- Elute mono-sulfated permethylated glycans with 100 μl of 2.5 mM NH4OAc in 50% ACN, and collect the eluent in the new Eppendorf tube.
- Elute di-sulfated permethylated glycans with 100 μl of 10 mM NH4OAc in 50% ACN, and collect the eluent in the new Eppendorf tube.
- Screen samples by MALDI-MS in the negative ion mode (the same as Procedure C).

Figure 2. The demonstration how to make self-packed amine tip. Step 1. Plug the yellow tip with filter paper by needle (shown in A). Step 2. Use 20 ml syringe as a plunger and put on a plastic hose on syringe to fit with yellow tip (shown in B). Step 3. Pack amine beads into the yellow tip by using syringe to wash out the solvent (shown in C).
- Prepare self-packed amine tip.
- Separation of sulfated permethylated glycans from non-sulfated ones by Oasis® MAX
- Equilibrate an Oasis® MAX cartridge with 3 ml of 95% ACN and 3 ml of 100 mM NH4OAc.
- Add 1 ml of 100 mM NH4OAc into the cartridge and load the neutralized reaction mixtures (Procedure A) and collect all the unbound samples in one glass tube. Reload the unbound fraction into the MAX cartridge.
- Wash the cartridge by 1 ml of 100 mM ammonium acetate, followed by 6 ml of water to remove as much salts as possible.
- Elute the non-sulfated permethylated glycans by 6 ml of 95% ACN, and collect the eluent in the new glass tube.
- Elute the mono-sulfated permthylated glycans by 6 ml of 1 mM NH4OAc in 80% ACN, and collect the eluent in the new glass tube.
- Elute permethylated glycans with more than one sulfate by 3 ml of 100 mM NH4OAc in 60% ACN, 20% methanol, and collect the eluent in the new glass tube.
- Dry samples by SpeedVac
Note: This mixed mode cartridge allows direct cleanup and sequential fractionation into non-sulfated, and sulfated glycans in one step.
- Further clean-up of permethylated glycans or concentration by ZipTipC18
- Transfer permethylated glycans (in ACN) into 0.5 ml or 1.5 ml Eppendorf tube and dry.
- Dissolve permethylated glycans in 20 μl of 10% ACN.
Note: To have better solubility of permethylated glycans in 10% ACN, use 2 μl of 100% ACN to re-dissolve and sequentially add 18 μl of water while constantly vortex the tube. - Equilibrate ZipTipC18 by 10 μl of 50% ACN, 0.1% TFA, then 10 μl of 0.1% TFA.
- Load sample by pipetting several times.
- Wash ZipTipC18 by 10 μl of 0.1% TFA, three times.
- Elute the glycans by 10 μl of 50% ACN, 0.1% TFA, or less volume, for collection into microtubes or direct spotting with matrix onto the MALDI target plate, followed by MS acquisition (Procedure C).
Data analysis
- All the MALDI-MS data are manually interpreted. In mammalian glycosylation, the common glycosyl residues are Hex, HexNAc, dHex (Fuc), Neu5Ac, and Neu5Gc, and the nominal masses of the permethylated forms are 204, 245, 174, 361, and 391 u, respectively. The accurate monoisotopic and average mass of different monosaccharide forms can be readily calculated or referring to Table I in Dell et al. (1994). For MALDI-MS of permethylated glycans in the negative ion mode, non-sulfated permethylated glycans do not normally afford any negative ion signal since all hydroxyl (OH) groups have been derivatized to O-methyl (OMe), and the carboxylic (COOH) group of sialic acids to methyl esters (COOMe).
- To calcgly charged m/z values of sulfated permethylated glycans in the negative ion mode, use the general formula:(CH3 + OCH3) corresponds to the respective non-reducing and reducing end masses, which is 46 u in nominal mass. For the O-glycans that were released from glycoproteins via reductive elimination, as well as any other pre-reduced N-glycans, their reducing end would be in an alditol state (annotated as -itol), and the corresponding nominal mass would be 62 u. (OSO3--OCH3) refers to the first sulfate carried, which corresponds to a 65 u mass increment, and contributing to the negative charge. Thereafter, for di- and multiply sulfated glycans, each additional sulfate group contributes to 88 u (65 + 23 u) mass increment from the mono-sulfated glycan due to the charge of second sulfate being counter balanced by a sodium. For examples in Figure 3, the [M-H]- molecular ion signal at m/z 1386 for a mono-sulfated NeuAc1Hex2HexNAc2-itol corresponds to [62 + 361 + 204 x 2 + 245 x 2 + 65], whereas the [M+Na-2H]- molecular ion signal at m/z 1562 for a di-sulfated Hex3HexNAc3-itol corresponds to [62 + 204 x 3 + 245 x 3 + 65 + 88].
- Mono-sulfated permethylated glycans, detected as [M-H]-
m/z = (CH3 + OCH3) + Σ glycosyl residue masses + (OSO3--OCH3) - Di-sulfated permethylated glycans, detected as [M+Na-2H]-
m/z = (CH3 + OCH3) + Σ glycosyl residue masses+ 2 x (OSO3--OCH3) +23
- Mono-sulfated permethylated glycans, detected as [M-H]-
- The signal corresponding to loss of sodium sulfite (-102 u, NaSO3) from di- and multiply sulfated glycans during MALDI ionization are often produced easily. Sometimes only these and not the ones retaining all sulfates were detected. It is also equivalent in mass to -14 u from the mono-sulfated species, and overlaps with any undermethylated form, if present (as shown in Figure 1.3A in Khoo and Yu (2010)).
Note: In practice, the relative signal intensity of monosodiated, di-sulfated permethylated glycans and those having lost the extra sodium sulfite varies from sample to sample and also somewhat depends on the MALDI-MS instrument conditions. In the case example shown in Figure 3B, the intensity of the di-sulfated O-glycan ion signal at m/z 1562 is much stronger than the signal at m/z 1460, but the reverse pattern has also been observed in other samples.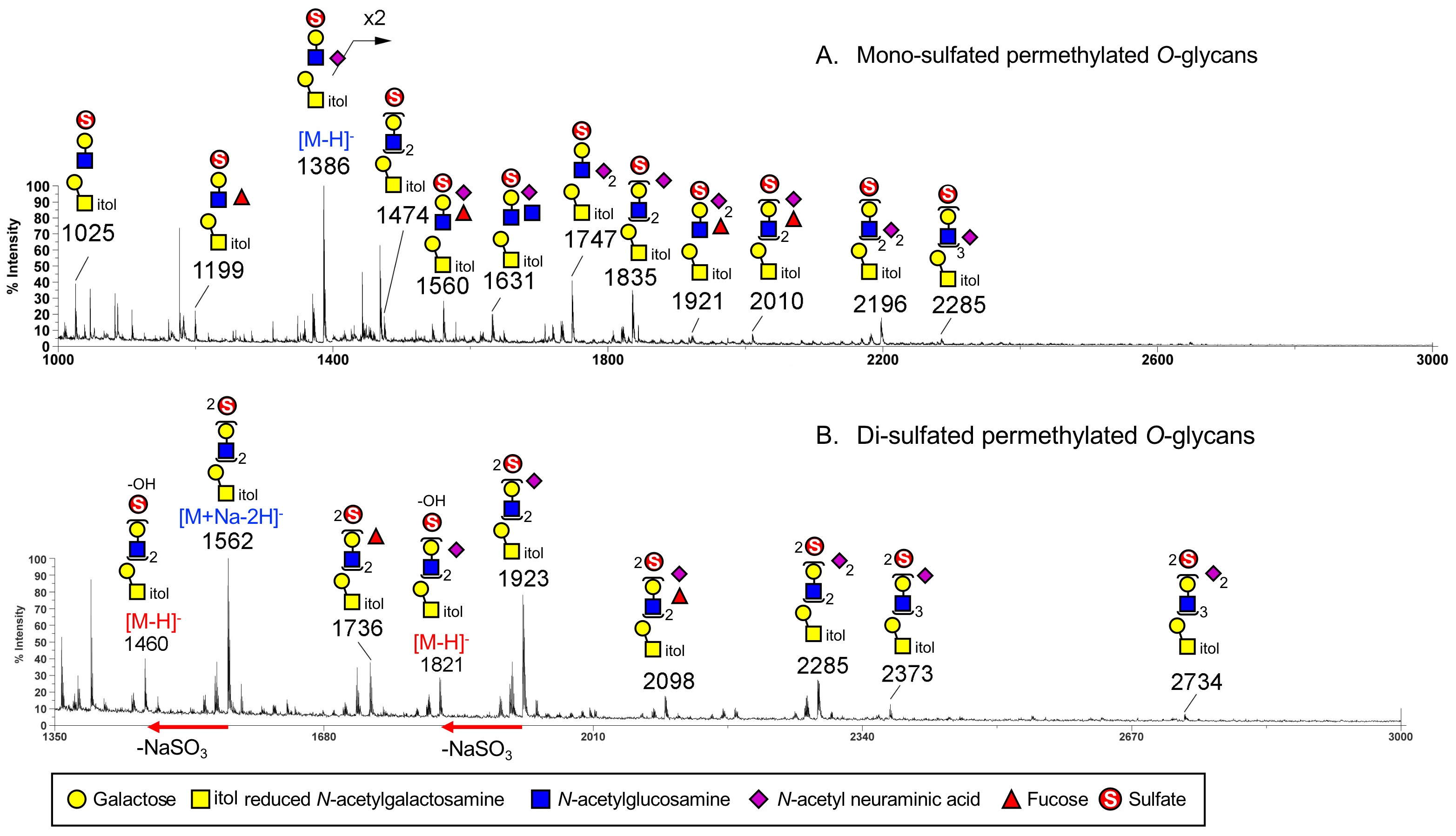
Figure 3. MALDI-MS profiles of mono-sulfated (A) and di-sulfated (B) permethylated O-glycans in the negative ion mode. The mono-sulfated and di-sulfated permethylated O-glycans were separated into different fractions by amine-beads, using the protocol described here. The mono-sulfated O-glycans were eluted in the earlier 2.5 mM NH4OAc in 50% ACN fraction (A) whereas the di-sulfated O-glycans in the 10 mM NH4OAc in 50% ACN fraction (B). The O-glycans shown here were reductively released from colonic cancer cell line SW480 transfected with GlcNAc6ST-2, as described in details in the original report (Yu et al., 2018).
Notes
The non-sulfated, sialylated glycans will co-elute with the sulfated glycans and be detected in the negative ion mode if the carboxylic group of their sialic acid remain non-methyl-esterified during the permethylation process. This undermethylation issue can be solved by re-permethylation of the recovered glycans. A prior clean-up of the native glycans by graphitized carbon column before permethylation may help achieving full permethylation. The detailed protocol was described in Khoo and Yu (2010).
Recipes
- 2.5% ACN (200 ml)
5 ml of 100% ACN
195 ml of ddH2O - 10% ACN (200 ml)
20 ml of 100% ACN
180 ml of ddH2O - 25% ACN (200 ml)
50 ml of 100% ACN
150 ml of ddH2O - 50% ACN (200 ml)
100 ml of 100% ACN
100 ml of ddH2O - 0.1% TFA (50 ml)
50 μl of TFA in 49.95 ml of ddH2O - 75% ACN, 0.1% TFA (50 ml)
37.5 ml of 100% ACN
12.45 ml of ddH2O
50 μl of TFA - 95% ACN (50 ml)
47.5 ml of 100% ACN
2.5 ml of ddH2O - 95% ACN, 0.1% FA (50 ml)
47.5 ml of 100% ACN
2.45 ml of ddH2O
50 μl of FA - 50% ACN, 0.1% FA (50 ml)
25 ml of 100% ACN
24.95 ml of ddH2O
50 μl of FA - 1 M ammonium acetate (NH4OAc)
Prepare 100 ml of 1 M ammonium acetate (NH4OAc) in deionized water as stock solution:
7.7 g NH4OAc, 100 ml of ddH2O - 50% ACN, 2.5 mM NH4OAc (50 ml)
25 ml of 100% ACN
0.125 ml of 1M NH4OAc
24.875 ml of ddH2O - 50% ACN, 10 mM NH4OAc (50 ml)
25 ml of 100% ACN
0.5 ml of 1 M NH4OAc
24.5 ml of ddH2O - 100 mM NH4OAc (200 ml)
20 ml of 1 M NH4OAc
180 ml of ddH2O - 80% ACN, 1 mM NH4OAc (200 ml)
160 ml of 100% ACN
0.2 ml of 1 M NH4OAc
39.8 ml of ddH2O - 60% ACN, 20% methanol, 100 mM NH4OAc (200 ml)
120 ml of 100% ACN
40 ml of methanol
20 ml of 1 M NH4OAc
20 ml of ddH2O
Acknowledgments
The sulfoglycomic workflow based on MALDI-MS analysis of permethylated glycans was originally developed by Yu et al. (2009), described in Khoo and Yu (2010), and further refined by introducing the MAX fractionation method, as described in Cheng et al. (2013). It was supported by Academia Sinica and Taiwan National Science Council grants to KKH and the NRPGM Core Facilities for Proteomics and Glycomics located at IBC, Academia Sinica.
Competing interests
The authors declare no conflicts of interest.
References
- Bochner, B. S., R. A. Alvarez, P. Mehta, N. V. Bovin, O. Blixt, J. R. White and R. L. Schnaar (2005). Glycan array screening reveals a candidate ligand for Siglec-8. J Biol Chem 280(6): 4307-4312.
- Cheng, C. W., Chou, C. C., Hsieh, H. W., Tu, Z., Lin, C. H., Nycholat, C., Fukuda, M. and Khoo, K. H. (2015). Efficient mapping of sulfated glycotopes by negative ion mode nanoLC-MS/MS-Based sulfoglycomic analysis of permethylated glycans. Anal Chem 87(12): 6380-6388.
- Cheng, P. F., Snovida, S., Ho, M. Y., Cheng, C. W., Wu, A. M. and Khoo, K. H. (2013). Increasing the depth of mass spectrometry-based glycomic coverage by additional dimensions of sulfoglycomics and target analysis of permethylated glycans. Anal Bioanal Chem 405(21): 6683-6695.
- Dell, A., Reason, A. J., Khoo, K. H., Panico, M., McDowell, R. A. and Morris, H. R. (1994). Mass spectrometry of carbohydrate-containing biopolymers. Methods Enzymol 230: 108-132.
- Harvey, D. J. (1993). Quantitative aspects of the matrix-assisted laser desorption mass spectrometry of complex oligosaccharides. Rapid Commun Mass Spectrom 7(7): 614-619.
- Hernandez Mir, G., Helin, J., Skarp, K. P., Cummings, R. D., Makitie, A., Renkonen, R. and Leppanen, A. (2009). Glycoforms of human endothelial CD34 that bind L-selectin carry sulfated sialyl Lewis x capped O- and N-glycans. Blood 114(3): 733-741.
- Hsiao, C. T., Wang, P. W., Chang, H. C., Chen, Y. Y., Wang, S. H., Chern, Y. and Khoo, K. H. (2017). Advancing a high throughput glycotope-centric glycomics workflow based on nanoLC-MS(2)-product dependent-MS(3) analysis of permethylated glycans. Mol Cell Proteomics 16(12): 2268-2280.
- Ideo, H., Seko, A., Ohkura, T., Matta, K. L. and Yamashita, K. (2002). High-affinity binding of recombinant human galectin-4 to SO3--->3Galβ1-->3GalNAc pyranoside. Glycobiology 12(3): 199-208.
- Jang-Lee, J., S. J. North, M. Sutton-Smith, D. Goldberg, M. Panico, H. Morris, S. Haslam and A. Dell (2006). Glycomic profiling of cells and tissues by mass spectrometry: fingerprinting and sequencing methodologies. Methods Enzymol 415: 59-86.
- Kawashima, H. (2006). Roles of sulfated glycans in lymphocyte homing. Biol Pharm Bull 29(12): 2343-2349.
- Khoo, K. H. and Yu, S. Y. (2010). Mass spectrometric analysis of sulfated N- and O-glycans. Methods Enzymol 478: 3-26.
- Kumagai, T., Katoh, T., Nix, D. B., Tiemeyer, M. and Aoki, K. (2013). In-gel β-elimination and aqueous-organic partition for improved O- and sulfoglycomics. Anal Chem 85(18): 8692-8699.
- Mitoma, J., Bao, X., Petryanik, B., Schaerli, P., Gauguet, J. M., Yu, S. Y., Kawashima, H., Saito, H., Ohtsubo, K., Marth, J. D., Khoo, K. H., von Andrian, U. H., Lowe, J. B. and Fukuda, M. (2007). Critical functions of N-glycans in L-selectin-mediated lymphocyte homing and recruitment. Nat Immunol 8(4): 409-418.
- Morelle, W. and Michalski, J. C. (2007). Analysis of protein glycosylation by mass spectrometry. Nat Protoc 2(7): 1585-1602.
- Patnode, M. L., Cheng, C. W., Chou, C. C., Singer, M. S., Elin, M. S., Uchimura, K., Crocker, P. R., Khoo, K. H. and Rosen, S. D. (2013). Galactose 6-O-sulfotransferases are not required for the generation of Siglec-F ligands in leukocytes or lung tissue. J Biol Chem 288(37): 26533-26545.
- Patnode, M. L., Yu, S. Y., Cheng, C. W., Ho, M. Y., Tegesjo, L., Sakuma, K., Uchimura, K., Khoo, K. H., Kannagi, R. and Rosen, S. D. (2013). KSGal6ST generates galactose-6-O-sulfate in high endothelial venules but does not contribute to L-selectin-dependent lymphocyte homing. Glycobiology 23(3): 381-394.
- Rosen, S. D. (2004). Ligands for L-selectin: homing, inflammation, and beyond. Annu Rev Immunol 22: 129-156.
- Shibata, T. K., Matsumura, F., Wang, P., Yu, S., Chou, C. C., Khoo, K. H., Kitayama, K., Akama, T. O., Sugihara, K., Kanayama, N., Kojima-Aikawa, K., Seeberger, P. H., Fukuda, M., Suzuki, A., Aoki, D. and Fukuda, M. N. (2012). Identification of mono- and disulfated N-acetyl-lactosaminyl Oligosaccharide structures as epitopes specifically recognized by humanized monoclonal antibody HMOCC-1 raised against ovarian cancer. J Biol Chem 287(9): 6592-6602.
- Wang, S. H., Tsai, C. M., Lin, K. I. and Khoo, K. H. (2013). Advanced mass spectrometry and chemical analyses reveal the presence of terminal disialyl motif on mouse B-cell glycoproteins. Glycobiology 23(6): 677-689.
- Yu, S. Y., Chang, L. Y., Cheng, C. W., Chou, C. C., Fukuda, M. N. and Khoo, K. H. (2013). Priming mass spectrometry-based sulfoglycomic mapping for identification of terminal sulfated lacdiNAc glycotope. Glycoconj J 30(2): 183-194.
- Yu, S. Y., Hsiao, C. T., Izawa, M., Yusa, A., Ishida, H., Nakamura, S., Yagi, H., Kannagi, R. and Khoo, K. H. (2018). Distinct substrate specificities of human GlcNAc-6-sulfotransferases revealed by mass spectrometry-based sulfoglycomic analysis. J Biol Chem 293(39): 15163-15177.
- Yu, S. Y., Wu, S. W., Hsiao, H. H. and Khoo, K. H. (2009). Enabling techniques and strategic workflow for sulfoglycomics based on mass spectrometry mapping and sequencing of permethylated sulfated glycans. Glycobiology 19(10): 1136-1149.
- Yu, S. Y., Cheng, C. W. and Khoo, K. H. (2020). Negative Ion Mode nanoLC-ESI-MS/MS Analyses of Permethylated Sulfated Glycans. Bio-protocol 10(10): e3618.
Article Information
Copyright
© 2020 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Readers should cite both the Bio-protocol article and the original research article where this protocol was used:
- Yu, S., Snovida, S. and Khoo, K. (2020). Permethylation and Microfractionation of Sulfated Glycans for MS Analysis. Bio-protocol 10(10): e3617. DOI: 10.21769/BioProtoc.3617.
- Yu, S. Y., Hsiao, C. T., Izawa, M., Yusa, A., Ishida, H., Nakamura, S., Yagi, H., Kannagi, R. and Khoo, K. H. (2018). Distinct substrate specificities of human GlcNAc-6-sulfotransferases revealed by mass spectrometry-based sulfoglycomic analysis. J Biol Chem 293(39): 15163-15177.
Category
Biochemistry > Carbohydrate > Polysaccharide
Biochemistry > Carbohydrate > Glycoprotein
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.