- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Tethered Chromosome Conformation Capture Sequencing in Triticeae: A Valuable Tool for Genome Assembly

Published: Vol 8, Iss 15, Aug 5, 2018 DOI: 10.21769/BioProtoc.2955 Views: 10121
Reviewed by: Rainer MelzerPooja VermaVinay Panwar
Original research article
The authors used this protocol in:

Apr 2017

Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Abstract
Chromosome conformation capture sequencing (Hi-C) is a powerful method to comprehensively interrogate the three-dimensional positioning of chromatin in the nucleus. The development of Hi-C can be traced back to successive increases in the resolution and throughput of chromosome conformation capture (3C) (Dekker et al., 2002). The basic workflow of 3C consists of (i) fixation of intact chromatin, usually by formaldehyde, (ii) cutting the fixed chromatin with a restriction enzyme, (iii) religation of sticky ends under diluted conditions to favor ligations between cross-linked fragments or those between random fragments and (iv) quantifying the number of ligations events between pairs of genomic loci (de Wit and de Laat, 2012). In the original 3C protocol, ligation frequency was measured by amplification of selected ligation junctions corresponding to a small number of genomic loci (‘one versus one’) through semi-quantitative PCR (Dekker et al., 2002). The chromosome conformation capture-on-chip (4C) and chromosome conformation capture carbon copy (5C) technologies then extended 3C to count ligation events in a ‘one versus many’ or ‘many versus many’ manner, respectively. Hi-C (Lieberman-Aiden et al., 2009) finally combined 3C with next-generation sequencing (Metzker, 2010). Here, before religation sticky ends are filled in with biotin-labeled nucleotide analogs to enrich for fragments with a ligation junction in a later step. The Hi-C libraries are then subjected to high-throughput sequencing and the resultant reads mapped to a reference genome, allowing the determination of contact probabilities in a ‘many versus many’ way with a resolution that is limited only by the distribution of restriction sites and the read depth. The first application of Hi-C was the elucidation of global chromatin folding principles in the human genome (Lieberman-Aiden et al., 2009). Similar efforts have since been carried out in other eukaryotic model species such as yeast (Duan et al., 2010), Drosophila (Sexton et al., 2012) and Arabidopsis (Grob et al., 2014; Wang et al., 2015; Liu et al., 2016). Other uses of Hi-C include the study of chromatin looping at high-resolution (Rao et al., 2014; Liu et al., 2016), of chromatin reorganization along the cell cycle (Naumova et al., 2013) and of differences in chromatin organization in mutant individuals (Feng et al., 2014). The tethered conformation capture protocol (TCC) (Kalhor et al., 2011) described here is a variant of the original Hi-C method (Lieberman-Aiden et al., 2009) and was adapted to Triticeae.
Keywords: Tethered conformation captureBackground
Any successful Hi-C experiment should reveal the distance-dependent decay of contact probabilities: the frequency by which two loci juxtapose in three-dimensional space is predominantly determined by their distance in the linear genome (Lieberman-Aiden et al., 2009). This observation motivated the application of Hi-C for physical mapping: the number of Hi-C links between pairs of contigs of a whole-genome shotgun assembly can serve as a proxy of the linear distance between them. This distance information can then be fed into graph algorithms to reconstruct the linear order of scaffolds along the chromosomes (Burton et al., 2013; Kaplan and Dekker, 2013). The three-dimensional proximity information obtained from our TCC approach was employed to order and orient BAC-based super scaffolds in the high-quality barley reference genome (Beier et al., 2017; Mascher et al., 2017). For the Emmer genome assembly, the TCC information was used in a similar manner to validate the scaffolds leading to chromosome-scale assemblies (pseudomolecules) (Avni et al., 2017).
TCC is a modified Hi-C approach in which key reactions (marking of the DNA ends and circularization) are performed on a solid phase rather than in solution (Kalhor et al., 2011). In fact, in human cells the tethering approach yielded improved signal-to-noise ratios, thus leading to a better mapping of low-frequency interactions (Kalhor et al., 2011). This observation prompted us to develop a similar protocol for Triticeae, which is based on the isolation of crosslinked nuclei from young leaves (Hövel et al., 2012) followed by TCC (Kalhor et al., 2011) (Figure 1). Briefly, the chromosome conformation is captured in native leaves by chemical crosslinking using formaldehyde (Hövel et al., 2012), which covalently connects proteins to DNA, as well as proteins to each other. Nuclei are purified, and cysteine residues of proteins contained in the chromatin are biotinylated (Kalhor et al., 2011). The DNA is digested with a restriction enzyme (HindIII) followed by immobilization at low density on streptavidin-coated beads via the crosslinked biotinylated proteins (Kalhor et al., 2011). DNA ends are filled-in, marked with biotin and circularized while tethered to the beads (Kalhor et al., 2011). The crosslinks are reversed, ligation junctions are affinity-purified and provided with adapters for Illumina sequencing to discover genuine pairs of intrachromosomal interaction sites, which were initially captured (Kalhor et al., 2011).
Chromosome conformation capture sequence data is analyzed in the context of genome sequence assemblies. If a chromosome-scale reference genome is available, Hi-C (or TCC) reads can be aligned to it and assigned to restriction fragment predicted in silico from the genome sequence. Subsequently, the number of Hi-C reads linking pairs of restriction pairs can be quantified and the counts be aggregated in larger genome windows (e.g., 1 Mb bins) to obtain estimates of the contact probabilities between pairs of genomic loci (Lieberman-Aiden et al., 2009). These contact probabilities can be used to investigate chromatin organization and its interaction with various biological parameters such as stage of the cell cycle (Naumova et al., 2013) or the epigenomic landscape (Zhou et al., 2013). Chromosome conformation capture sequencing can also be used to reconstruct contiguous, chromosome-scale genomic reference sequences from fragmented sequence assembly. The Hi-C/TCC reads are mapped to a sequence assembly composed of unordered scaffolds, and links between scaffolds are counted and used as a measure of genomic distance. The closer two sequence scaffolds are to each other in the linear genome, the higher the number of Hi-C read pairs linking them. This distance information can then be used to derive a linear order of sequence scaffolds and orient adjacent scaffolds relative to each other (Burton et al., 2013; Kaplan and Dekker, 2013). These approaches have been applied in the sequence assembly of insect (Dudchenko et al., 2017) and Triticeae genomes (Avni et al., 2017; Mascher et al., 2017).
Figure 1. Schematic overview of TCC for Triticeae. Leaves of seven days old plants (I) are harvested for chemical crosslinking of the chromatin (II). The chromatin structure is captured by the formation of covalent bonds between proteins (grey ovals) and between proteins and DNA (line). The orange and light blue segments exemplify two HindIII fragments located on a chromosome. Nuclei are purified (III), and proteins are biotinylated (IV, green pentagon) for subsequent tethering of the complexes to a solid phase. DNA is digested (scissors) using the restriction enzyme HindIII (V) and tethered at low density to streptavidin-coated beads (VI, purple). Ends are marked and filled-in using biotin-14-dCTP (VII, red star). By including dGTPαS in the fill-in reaction, a phosphorothioate bond is introduced. Thereby DNA is guarded against Exonuclease III digestion, which is used at a later step to remove biotinylated nucleotides from non-ligated ends (not shown). Filled-in HindIII sites are ligated ‘on-bead’ (VIII), thereby newly creating NheI restriction sites, which are indicating TCC ligation events. The crosslinking is reversed. DNA is purified (IX) and treated with Exonuclease III (not shown) prior to fragmentation (X). The position of primers (grey triangles) used for controlling the marking and ligation of ends (3C control) is indicated (IX). Ends of the fragmented DNA are repaired and tailed with ‘A’ (not shown). DNA fragments with genuine ligation junctions are affinity-purified based on the incorporated biotin (XI) and provided with Illumina adapters (XII, grey lines) for paired-end sequencing. Ligation products are PCR-amplified (XII; arrowheads: position of primer), size-selected (XIII) and quality controlled (QC). The junctions are revealed by paired-end (PE) sequencing using an Illumina HiSeq2500 device (XIV) followed by bioinformatic analysis (XV). The estimated hands-on time is indicated in days (d). The operating period of the Illumina HiSeq2500 instrument is given for 2x 100 cycles sequencing and depends on the chemistry and type of flowcell (days in brackets). The flowchart is adapted from (Kalhor et al., 2011).
Materials and Reagents
Notes:
- We tested all components in independent experiments. However, this list does not imply that alternative products from other manufacturers cannot perform just as well.
- The use of the published equipment and chemistry is not indicating any competing interest.
- Compost soil (multiple vendors)
- Pots (16 cm diameter) (multiple vendors)
- Aluminum foil (multiple vendors)
- 10 µl Sapphire Low Retention Filtertips (Greiner Bio One International, catalog number: 771265 )
- 200 µl Sapphire Low Retention Filtertips (Greiner Bio One International, catalog number: 737265 )
- 1,000 µl Sapphire Low Retention Filtertips (Greiner Bio One International, catalog number: 750265 )
- 0.2 µm filter, Millex GP (Merck, catalog number: SLGP033RS )
- 10 ml and 50 ml syringes (multiple vendors)
- 50 ml Falcon tubes (Corning, catalog number: 352070 )
- Racks for 50 ml Falcon tubes fitting into the desiccator (multiple vendors)
- Disposable 10 ml and 25 ml plastic pipettes (multiple vendors)
- Pipette fillers (BRAND, catalog number: 25315 )
- Paper towels (multiple vendors)
- Appropriate protective gear for working with liquid nitrogen (multiple vendors)
- Dewar vessel for transport and storage of liquid nitrogen (multiple vendors)
- Miracloth (Merck, catalog number: 475855-1R )
- Sefar Nitex 03-55/32 (55 µm mesh opening, 32% open area) (Stefan Kastenmüller GmbH)
- Tubes (1.5 ml and 2.0 ml; SARSTEDT, catalog numbers: 72.690.001 and 72.695.500 )
- Microscopic slides (76 x 26 x 1 mm) (multiple vendors)
- Cover slips (multiple vendors)
- Cell counting chamber (Neubauer chamber) and glass cover (Celeromics)
- Needle (0.9 x 40 mm, HSW FINE-JECT) (Henke-Sass, Wolf, catalog number: 8300025263 )
- Slide-A-LyzerTM Dialysis cassette (0.5-3 ml, cutoff 20 kDa) with float buoys (Thermo Fisher Scientific, catalog number: 66003 )
- Parafilm (BRAND, catalog number: 701605 )
- Magnetic Particle Concentrator (MPC) for 1.5 ml tubes (DynaMag-2 Magnet) (Thermo Fisher Scientific, catalog number: 12321D )
- 15 ml conical tubes (Corning, catalog number: 352196 )
- Rubber strap or tape
- DynaMag-96 Side Skirted Magnetic Particle Concentrator (MPC96) (Thermo Fisher Scientific, catalog number: 12027 )
- PCR-tubes, Sapphire (Greiner Bio One International, catalog number: 652250 )
- Snap-cap microTUBEs (with AFA-fiber and pre-split septum) (Covaris, catalog number: 520045 )
- Disposable scalpels (multiple vendors)
- Polystyrene plugs (self-made) of 0.5 to 1.0 cm thickness fitting snugly into the 50 ml plastic tubes
- Seeds (provided by the experimenter)
- HEPES sodium salt (4-(2-Hydroxyethyl) piperazine-1-ethanesulfonic acid sodium salt) (Carl Roth, catalog number: 7020.2 )
- Concentrated HCl (Carl Roth, catalog number: 4625.1 )
- Sucrose (Carl Roth, catalog number: 9097.1 )
- MgCl2•6H2O (Sigma-Aldrich, catalog number: M2670 )
- KCl (Honeywell, Fluka, catalog number: 60130 )
- Triton X-100 (Carl Roth, catalog number: 3051.3 )
- 100% glycerol (Carl Roth, catalog number: 3783.1 )
- Phenylmethylsulfonyl fluoride (PMSF) (Sigma-Aldrich, catalog number: 93482 )
- 2-mercaptoethanol (14.3 M) (Carl Roth, catalog number: 4227.1 )
- 37% (w/v) formaldehyde solution (containing 10-15% methanol as stabilizer) (Sigma-Aldrich, catalog number: 252549 )
- Glycine (Carl Roth, catalog number: 3790.1 )
- Liquid nitrogen
- Aprotinin (Sigma-Aldrich, catalog number: 10820 )
- Leupeptin (Sigma-Aldrich, catalog number: L2884 )
- Pepstatin (Sigma-Aldrich, catalog number: P5318 )
- VECTASHIELD mounting medium with DAPI (VECTOR Laboratories, catalog number: H-1200 )
- SDS (Sodium Dodecyl Sulfate) (Serva, catalog number: 20765 )
- Tris (hydroxymethyl) aminomethane (Tris base) (Carl Roth, catalog number: 5429.3 )
- NaCl (Carl Roth, catalog number: 9265.2 )
- EZlink Iodoacetyl-PEG2-Biotin (IPB) (Thermo Fisher Scientific, catalog number: 21334 )
- 1,4-Dithiothreitol (DTT) (Carl Roth, catalog number: 6908.1 )
- HindIII (100 U/µl) (New England BioLabs, catalog number: R0104T )
- NheI (10 U/µl) (Thermo Fisher Scientific, catalog number: ER0971 )
- MyOne Streptavidin T1 beads (Thermo Fisher Scientific, catalog number: 65601 )
- Na2HPO4 (Carl Roth, catalog number: P030.2 )
- KH2PO4 (Carl Roth, catalog number: 3904.1 )
- 100 mM dATP (Thermo Fisher Scientific, catalog number: R0142 )
- 100 mM dTTP (Thermo Fisher Scientific, catalog number: R0172 )
- 2'-Deoxyguanosine-5'-(α-thio)-triphosphate, Sodium salt (dGTPαS), 1:1 Mixture of Rp and Sp isomers (Jena Bioscience, catalog number: NU-424S )
- Biotin-14-dCTP (Thermo Fisher Scientific, catalog number: 19518018 )
- Klenow DNA polymerase, large fragment (10 U/µl) (Thermo Fisher Scientific, catalog number: EP0052 )
- UltraPure BSA (50 mg/ml) (Thermo Fisher Scientific, catalog number: AM2616 )
- T4 DNA ligase (5 U/µl) with 10x T4 DNA Ligation buffer and 50% PEG 4000 (Thermo Fisher Scientific, catalog number: EL0011 )
- RNase A (DNase-free) (MACHEREY-NAGEL, catalog number: 740505 )
- Proteinase K (Thermo Fisher Scientific, catalog number: 25530015 )
- Phenol:CHCl3:isoamyl alcohol (25:24:1) (Carl Roth, catalog number: A156.2 )
- 8-Hydroxyquinoline (Sigma-Aldrich, catalog number: H6878 )
- Isoamyl alcohol (Carl Roth, catalog number: 8930.1 )
- CHCl3 (Carl Roth, catalog number: 3313.2 )
- Glycogen (Thermo Fisher Scientific, catalog number: R0561 )
- 100% ethanol (Carl Roth, catalog number: 9065.4 )
- Q5 Hot Start High-Fidelity DNA Polymerase (2 U/µl) and 5x Q5 Hot Start buffer (New England BioLabs, catalog number: M0493S )
- Primer for biotin incorporation control (barley):
Primer 1 (5'- ATCTTCATGCGAGGCAGAGT-3')
Primer 2 (5'- ACCGTTGAACCATCTTCAGG-3')
Note: The primers are HPLC purified, 0.2 μmol synthesis scale, dissolved in H2O to 100 µM; contact e.g., Sigma-Aldrich for synthesis. - dNTP Set (100 mM solutions) (Thermo Fisher Scientific, catalog number: R0181 )
- dNTP Mix (25 mM each) (Thermo Fisher Scientific, catalog number: R1121 )
- MinElute Gel Extraction Kit (QIAGEN, catalog number: 28606 )
- Bromophenol Blue (Carl Roth, catalog number: A512.2 )
- 10x NEBuffer 1 buffer (New England BioLabs, catalog number: B7001S )
- UltraPure Agarose (Thermo Fisher Scientific, InvitrogenTM, catalog number: 16500500 )
- Exonuclease III (100 U/µl) from E. coli (New England BioLabs, catalog number: M0206S )
- AMPure XP beads (Beckman Coulter, catalog number: A63881 )
- 10x Tango (Thermo Fisher Scientific, catalog number: BY5 )
- 100 mM ATP (Thermo Fisher Scientific, catalog number: R0441 )
- T4 DNA polymerase (5 U/µl) (Thermo Fisher Scientific, catalog number: EP0062 )
- T4 polynucleotide kinase (10 U/µl) (Thermo Fisher Scientific, catalog number: EK0031 )
- Klenow DNA polymerase, large fragment (10U/µl) (Thermo Fisher Scientific, catalog number: EP0052 )
- Klenow fragment, 3'→5' exo- (5 U/µl) (New England BioLabs, catalog number: M0212L )
- Dynabeads MyOne Streptavidin C1 (Thermo Fisher Scientific, catalog number: 65001 )
- DNA LoBind tubes (1.5 ml) (Eppendorf, catalog number: 0030108051 )
- TruSeq DNA Single Indexes Set A (Illumina, catalog number: 20015960 )
- Library amplification primer:
Primer 3 (5'-AATGATACGGCGACCACCGAGAT-3')
Primer 4 (5'-CAAGCAGAAGACGGCATACGA -3')
Note: The primers are HPLC purified, 0.2 μmol synthesis scale, dissolved in H2O to 100 µM; contact e.g., Sigma-Aldrich for synthesis. - SYBR-Gold (Thermo Fisher Scientific, catalog number: S11494 )
- GeneRuler 50 bp DNA ladder (Thermo Fisher Scientific, catalog number: SM0373 )
- Glacial acetic acid (Carl Roth, catalog number: 3738.5 )
- QIAquick PCR Purification Kit (QIAGEN, catalog number: 28106 )
- 100% isopropanol (Carl Roth, catalog number: 6752.4 )
- 1 M Tris-HCl, pH 8.0 (100 ml)
- 1 M Tris-HCl, pH 7.4 (100 ml)
- 1 M Tris-HCl, pH 7.5 (100 ml)
- 0.5 M EDTA, pH 8.0 (100 ml)
- 1 M DTT (20 ml)
- Phenol:CHCl3:isoamyl alcohol (25:24:1) with 0.1% 8-Hydroxyquinoline
- CHCl3:isoamyl alcohol (24:1)
- 1 M MgCl2
- 1 M HEPES, pH 8.0 (see Recipes)
- 2 M sucrose (see Recipes)
- 1 M KCl (see Recipes)
- 10% (v/v) Triton X-100 (see Recipes)
- Nuclei Isolation Buffer (NIBF) with formaldehyde (see Recipes)
- 2 M glycine (see Recipes)
- 1 mg/ml Aprotinin (see Recipes)
- 1 mg/ml Leupeptin (see Recipes)
- 1 mg/ml Pepstatin (see Recipes)
- Nuclei Isolation Buffer with protease inhibitors (NIBP) (see Recipes)
- Sucrose cushion (see Recipes)
- Nuclei Isolation Buffer with 1.5 M sucrose (NIBS) (see Recipes)
- 2% (w/v) SDS (see Recipes)
- 4 M NaCl (see Recipes)
- Wash buffer 1 (see Recipes)
- 25 mM IPB (see Recipes)
- 10x NEBuffer 2 (see Recipes)
- 1x NEBuffer 2 (see Recipes)
- HindIII (10 U/ µl) (see Recipes)
- Dialysis buffer (TE buffer) (see Recipes)
- Phosphate Buffered Saline with Tween 20, pH 7.4 (PBST) (see Recipes)
- 25 mM neutralized IPB (see Recipes)
- 10 mM dATP (see Recipes)
- 10 mM dTTP (see Recipes)
- Wash buffer 2 (see Recipes)
- Wash buffer 3 (see Recipes)
- 10x ligation buffer TCC (see Recipes)
- 10 mg/ml BSA (see Recipes)
- Extraction buffer (see Recipes)
- 25 mg/ml RNase A (see Recipes)
- 20 mg/ml Proteinase K (see Recipes)
- 5 mg/ml glycogen (see Recipes)
- 80% ethanol (see Recipes)
- 70% ethanol (see Recipes)
- 3 M sodium acetate, pH 5.2 (see Recipes)
- EB (see Recipes)
- EBT (see Recipes)
- Primer 1 + 2 (10 µM each) (see Recipes)
- 10 mM dNTP mix (see Recipes)
- 2.5 mM dNTP mix (see Recipes)
- TLE (see Recipes)
- Tween Wash Buffer (TWB) (see Recipes)
- 2x Binding Buffer (2x BB) (see Recipes)
- 1x Binding Buffer (1x BB) (see Recipes)
- 1x ligation buffer (see Recipes)
- Primer 3 + 4 (10 µM each) (see Recipes)
- 6x loading dye (see Recipes)
- 50x TAE (see Recipes)
Equipment
- Scissors (multiple vendors)
- Ice bucket (multiple vendors)
- Borosilicate glass bottles (100 ml) (Laborbedarfshop, catalog number: GT00205 )
- Borosilicate glass bottles (250 ml) (Laborbedarfshop, catalog number: GT00206 )
- Borosilicate glass bottles (500 ml) (Laborbedarfshop, catalog number: GT00207 )
- Borosilicate glass bottles (1,000 ml) (Laborbedarfshop, catalog number: GT00208 )
- Borosilicate Erlenmeyer flasks (500 ml) (Laborbedarfshop, catalog number: GT00153 )
- P1, 10 µl Finnpipette (Thermo Fisher Scientific, catalog number: 4641040N )
- P10, 100 µl Finnpipette (Thermo Fisher Scientific, catalog number: 4641070N )
- P20, 200 µl Finnpipette (Thermo Fisher Scientific, catalog number: 4641080N )
- P100, 1000 µl Finnpipette (Thermo Fisher Scientific, catalog number: 4641100N )
- Mortar with pestle with a rough surface for grinding (about 10 cm diameter) (multiple vendors)
- Metal spoon (multiple vendors)
- Funnels fitting into 50 ml tubes (multiple vendors)
- Sieve or tea strainer (about 8 cm diameter) (multiple vendors)
- Greenhouse equipped with automatic shading and supplementary light (sodium halogen lamps)
- pH meter (multiple vendors)
- Water bath (65 °C) (multiple vendors)
- -20 °C freezer (multiple vendors)
- -80 °C freezer (multiple vendors)
- Vortex (multiple vendors)
- Measuring cylinder 100 ml, 1 L (multiple vendors)
- Autoclave (multiple vendors)
- Analytical laboratory balance 'Quintix' (0,1 mg to 120 g) (Sartorius, catalog number: Quintix® 124-1S )
- Precision laboratory balance 'Cubis' (10 mg to 2.2 kg) (Sartorius, catalog number: MSE2203P-000-DR )
- Ice machine (multiple vendors)
- Desiccator (multiple vendors)
- Vacuum pump with manometer, condensation trap and tubing to connect desiccator to vacuum pump (multiple vendors)
- Fume hood
Note: All manipulations involving the NIBF buffer must be performed inside a fume hood. Follow the safety regulations of your laboratory during the manipulations and for the waste disposal. - Heraeus Multifuge 4KR centrifuge for 50 ml tubes (3,000 x g required) (Thermo Fisher Scientific, model: HeraeusTM Multifuge 4KR , catalog number: 75004461)
- Heraeus Fresco 21 centrifuge for Eppendorf tubes (16,000 x g required) (Thermo Fisher Scientific, model: HeraeusTM FrescoTM 21 , catalog number: 75002426)
- Epifluorescence microscope BX61 (Olympus) equipped with a cooled CCD camera (Hamamatsu Orca ER) (see Note 1) (Olympus, model: BX61 )
- Incubator cabinet (37 °C) (Memmert, model: Model 600 , catalog number: D06062)
- Rocking platform (Heidolph Instruments, model: Titramax 1000, catalog number: 544-12200-00 )
- Eppendorf ThermoMixer C (Eppendorf, model: ThermoMixer® C , catalog number: 5382000015) with a Smartblock for 1.5 ml tubes (Eppendorf, catalog number: 5360000038 )
- 1 L beaker glass (multiple vendors)
- Magnetic stir bar and magnetic stirrer (multiple vendors)
- Tube rotator (NeoLab, catalog number: 7-0045 )
- Incubator cabinet (16 °C) (GFL-Gesellschaft für Labortechnik, catalog number: 3032 ) placed in a cold room at 4 °C
- Qubit 2.0 (or Qubit 4) fluorometer with assay tubes, dsDNA HS Assay and dsDNA BR Assay (Thermo Fisher Scientific, model: Qubit 4, catalog number: Q33227 )
- Thermocycler (multiple vendors)
- Microwave (multiple vendors)
- Agarose gel electrophoresis equipment and accessories [microwave, tray (15 x 15 cm), combs, power supply, electrophoresis buffer, agarose, etc.] (multiple vendors)
- Covaris S220 AFA Ultrasonicator (Covaris) and associated equipment [snap-cap microTUBEs (with AFA-fiber and pre-split septum), chiller, software, computer (see Note 1)]
- Dark Reader blue light transilluminator (Clare Chemical Research, catalog number: DR46B )
- Agilent 2100 Electrophoresis Bioanalyzer or Agilent 4200 TapeStation System (Agilent Technologies, model: Agilent 2100 ) including accessories and consumables (see Note 1)
- GenPure Pro UV/UF (Thermo Fisher Scientific, catalog number: 50131950 )
Software
Software and computer hardware:
To analyze Hi-C/TCC sequence data, a computer server running a Unix operating system (e.g., Linux, Solaris, MacOS; see Note 1) or access to a cloud-computing system (e.g., CyVerse, http://www.cyverse.org) is required. Common UNIX command line tools (such as) need to be available. To accelerate CPU-intensive steps such as read alignment, access to a multi-core machine (> 16 CPU cores) is recommended. Depending on the number of samples to be analyzed, hard disk storage space needs to be allocated.
The following bioinformatics software need to be installed to carry out the primary data analysis described below:
- Casava, http://support.illumina.com/sequencing/sequencing_software/casava.html
- Cutadapt, http://cutadapt.readthedocs.io
- BWA-MEM, https://github.com/lh3/bwa
- SAMtools, http://www.htslib.org/download/
- BEDtools, http://bedtools.readthedocs.io
Procedure
Notes:
- All procedures can be performed at room temperature unless specified otherwise.
- In order to avoid contaminations of the sequencing libraries, filter tips and gloves should be used.
- Surfaces should be cleaned for DNA removal with commercial products at regular intervals.
- Lab space and instruments for handling samples pre- and post-PCR must be separated physically.
- Although the protocol worked well (> 40 independent experiments), the authors take no liability for the success of the experiments conducted by the reader.
- Plant growth and harvesting
- About 100 barley seeds are planted in two pots (16 cm diameter) filled with compost soil. Water thoroughly, and grow the plants for 7 days in a greenhouse (sodium halogen lamps, light period of 16 h, night: 18 °C and day: 21 °C). As an example, growth conditions for barley are given in (Zimmermann et al., 2006). Other plant species may require different conditions. Adapt the plant cultivation accordingly. Use young tissues (unexpanded cells) with a favorable content of nuclei.
- Grow sufficient plants to obtain about 2.4 g of leaves.
- Harvest the leaves with scissors. Wrap the leaves in aluminum foil and store on ice for the transport to the lab until ready for next step.
- Chemical crosslinking
- Trim the leaves to 0.5-1.0 cm long segments and transfer about 0.8 g tissue into a 50 ml tube. Prepare 3 tubes and store on ice.
- Add inside the fume hood 15 ml cold NIBF to each tube. Push a polystyrene plug down to the liquid level for keeping the leaves submersed in the buffer during vacuum infiltration (Hövel et al., 2012) (Figure 2).

Figure 2. Tube setup during vacuum infiltration. A polystyrene plug fitting snugly to the tube is pushed down to the liquid surface area in order to ensure submersion of the leaf segments during the infiltration. - Place tubes in a desiccator and infiltrate the NIBF (150 mbar). Cut the vacuum abruptly after 5, 10 and 15 min to help the entering of the NIBF into the leaf segments. Continue with the infiltration for a total time of 1 h. The green color of segments will be darker after the infiltration (Figure 3) due to the penetration of the fixative into the intercellular space (Hövel et al., 2012) (see Note 2).

Figure 3. Leaf color change. The image is taken from a typical leaf segment before (A) and after (B) vacuum infiltration with NIBF buffer (fixative). - Cut the vacuum, remove the polystyrene plugs and add 2.0 ml of 2 M glycine to stop crosslinking (Hövel et al., 2012). Mix the sample thoroughly by pipetting up and down using a disposable 25 ml plastic pipette. Insert the plugs and apply vacuum (150 mbar) for 5 min.
- Dispose the plugs and decant the liquid into a waste container. Use a sieve to keep the leaf segments in the 50 ml tube (Figure 4). Wash the leaves three times by adding 40 ml purified water per tube.
- Dry the segments of each tube well with paper towel and store them on ice until grinding (Figure 4).

Figure 4. Washing and drying of leaf segments after chemical crosslinking. A. Segments are washed with purified water. A sieve is used to withhold the segments in the tube during decantation. B. Leaf segments from each tube (about 0.8 g) are dried thoroughly on a paper towel to facilitate subsequent grinding in liquid nitrogen.
- Cell disruption and nuclei isolation
- Precool mortar and pestle in liquid nitrogen (see Note 3). Grind about 0.8 g leaves to a fine powder. Use a metal spoon (precooled in liquid nitrogen) to transfer the powder into a pre-cooled 50 ml plastic tube (Figure 5A). Close the tube with a lid, which is punctured several times to ensure pressure balance. Store the material immediately at -80 °C. Prepare the remaining 2 tubes accordingly. Process the samples for nuclei isolation within 1 week.
- Place the 3 tubes containing the powder on ice after removing the tubes from the -80° freezer and add 10 ml NIBP (Hövel et al., 2012) to each tube using a 25 ml plastic pipette. Mix carefully with a pipette to resuspend frozen clumps completely.
- In order to prevent filters from clogging, distribute the suspension evenly into four funnels prepared in advance (as described below).
- Filter the suspension at 4 °C through Miracloth and Sefar Nitex placed in a funnel as described (Hövel et al., 2012). Miracloth should face the plant material and Sefar Nitex the funnel surface. Use gravity flow only to avoid contamination with cell debris. Collect the filtrate in a 50 ml plastic tube (Figure 5B).

Figure 5. Cell disruption and filtering of nuclei. A. Leaf segments are ground in liquid nitrogen to a fine powder. The material is transferred into a 50 ml tube using a metal spoon and immediately stored at -80 °C. B. For the isolation of nuclei, the powder is resuspended in 10 ml NIBP and filtered through Miracloth and Sefar Nitex. The extract is collected in a clean 50 ml tube for subsequent centrifugation. - Spin the four filtrates in a pre-cooled swing-out centrifuge (3,000 x g, 15 min, 4 °C). Discard the supernatant using a 10 ml plastic pipette and resuspend each pale green pellet in 1 ml ice-cold NIBS (Hövel et al., 2012).
- Transfer the suspension to four 1.5 ml tubes and spin in a pre-cooled centrifuge (1,900 x g, 5 min, 4 °C). Discard the supernatant and resuspend the pellets in 300 µl ice-cold NIBS.
- Prepare four 2.0 ml tubes each containing 1.5 ml ice-cold sucrose cushion and carefully layer 300 µl of the suspension on top essentially as described (Abdalla et al., 2009) (Figure 6). Spin the four tubes in a pre-cooled centrifuge (16,000 x g, 1 h, 4 °C). Remove the supernatant completely and avoid contaminating the colorless nuclei pellet with the pale green top layer.

Figure 6. Sucrose cushion. The pale green nuclei extract (e) is layered on top of a sucrose cushion (c) and centrifuged in a 2.0 ml tube for further purification. - Resuspend each of the four nuclei pellets in 100 µl ice-cold NIBS. Pool two nuclei suspensions (200 µl) in a 1.5 ml tube. Place the two pools on ice and perform a microscopic quality check.
- Microscopic quality and quantity check of the nuclei (optional)
Note: The microscopic check might be skipped, if nuclei are purified in a routine manner, or if an epifluorescence microscope is not available.
- Stain 7 µl nuclei suspension with 7 µl VECTASHIELD mounting medium with DAPI for the integrity examination (Hövel et al., 2012). Apply the stained sample to a microscopic slide and add the coverslip. Analyze the nuclei using the epifluorescence microscope (100x objective, 1,000-fold magnification, DAPI-filter, absorption 358 nm, emission 461 nm). Intact nuclei appear round or oval and show sharp contours (Figure 7).

Figure 7. Purified barley nuclei. The integrity of barley nuclei following sucrose cushion centrifugation is examined using epifluorescence microscopy (1,000-fold magnification) and staining with DAPI. The scale bar corresponds to 5 µm. - Stain another aliquot of the nuclei suspension with VECTASHIELD mounting medium with DAPI for quantification. Count the nuclei using a cell counting chamber (Neubauer chamber) and the epifluorescence microscope (20x objective, 200-fold magnification, DAPI-filter, absorption 358 nm, emission 461 nm). Determine the concentration of the nuclei and continue the TCC library construction with approximately 107 nuclei.
- Stain 7 µl nuclei suspension with 7 µl VECTASHIELD mounting medium with DAPI for the integrity examination (Hövel et al., 2012). Apply the stained sample to a microscopic slide and add the coverslip. Analyze the nuclei using the epifluorescence microscope (100x objective, 1,000-fold magnification, DAPI-filter, absorption 358 nm, emission 461 nm). Intact nuclei appear round or oval and show sharp contours (Figure 7).
- Chromatin biotinylation
- Spin the two tubes with the nuclei suspension in a pre-cooled centrifuge (1,900 x g, 5 min, 4 °C) and discard the supernatant. Resuspend each pellet in 900 µl ice-cold wash buffer 1 (Kalhor et al., 2011). Spin the sample in a pre-cooled centrifuge (1,900 x g, 5 min, 4 °C) and discard the supernatant.
- Repeat the washing step with wash buffer 1.
- Resuspend each pellet with wash buffer 1. Each tube should contain a volume of 125 µl sample (total volume of the entire nuclei preparation: 250 µl).
- Add 48 µl 2% SDS per tube and incubate in a pre-warmed water bath (65 °C, 10 min) to solubilize the crosslinked chromatin (Kalhor et al., 2011). Cool samples to room temperature.
- Add 53 µl 25 mM IPB per tube and agitate in a light-tight container (1 h) to biotinylate the cysteine residues (Kalhor et al., 2011).
- For neutralization of SDS add 650 µl 1x NEBuffer 2 per tube and incubate on ice for 5 min (Kalhor et al., 2011).
- Add 113 µl 10% Triton X-100 per tube, mix, incubate on ice (10 min) and at 37 °C (10 min) (Kalhor et al., 2011).
- The total reaction volume is 1,975 µl (2 tubes with 989 µl each).
- HindIII digestion and dialysis
- Add 50 µl 10x NEBuffer 2, 2.5 µl 1 M DTT, 215 µl water and 10 µl HindIII (100 U/µl) to each tube (Kalhor et al., 2011).
- The total volume per tube is 1,267 µl.
- Incubate the two tubes horizontally in an incubator cabinet (37 °C, overnight, gentle agitation on a rocking platform).
- Pool the two digestions and inject the sample (about 2.5 ml) into one pre-wetted Slide-A-LyzerTM Dialysis cassette (20 kDa, 3 ml) according to the manufacturer’s instructions. Inject slowly to avoid shearing of the DNA (Figure 8). Aspirate most of the remaining air in the cassette, fill 1 L dialysis buffer into a beaker glass, add a magnetic stir bar and dialyze the cassette inserted into a float buoy with slow agitation for 3 h, thereby removing excess IPB from the biotinylation reaction (Kalhor et al., 2011).

Figure 8. Dialysis. After the HindIII digestion, the sample (about 2.5 ml) is injected slowly into a pre-wetted Slide-A-LyzerTM Dialysis cassette. Prior to dialysis, most of the air is aspirated from the inside. The cassette is inserted into a float buoy and placed into dialysis buffer for a total of 4 h. - Discard the buffer and dialyze for 1 h in 1 L fresh dialysis buffer.
- Collect the DNA as described by the manufacturer of the dialysis cassette. Draw the sample slowly into the syringe to avoid shearing.
- Transfer the sample evenly into two 2.0 ml tubes.
- Tethering
- Prepare MyOne Streptavidin T1 beads (Kalhor et al., 2011). Resuspend the beads by gentle vortexing and dispense 200 µl in each of two 1.5 ml tubes (see Note 4).
- Place the tubes into a MPC, wait for 3 min and discard the supernatant.
- Remove the tubes from the MPC, add 1 ml PBST per tube and resuspend the beads by gentle vortexing. Place the tubes in the MPC for 3 min and discard the supernatant.
- Repeat the washing step with PBST twice.
- Resuspend the beads in each tube in 1 ml PBST (total volume 2 ml).
- Distribute the dialyzed sample equally into five 1.5 ml tubes and add PBST to a final volume of 500 µl per tube. Label 4 tubes ‘TCC’ and one tube ‘3C’.
- Add to each tube 400 µl washed MyOne Streptavidin T1 beads (final volume 900 µl per tube) and place the samples in a tube rotator for 30 min.
- During the affinity binding prepare the 25 mM neutralized IPB. Add 7 µl 25 mM neutralized IPB to each of the 5 tubes and place the samples in a tube rotator for 15 min (see Note 5).
- Marking DNA ends and circularization
- Place the five tubes into an MPC, wait for 2 min and discard the supernatant.
- Remove the tubes from the MPC, add 600 µl PBST (Kalhor et al., 2011) per tube and resuspend the beads by gentle vortexing. Place the tubes in the MPC for 2 min and discard the supernatant.
- Repeat the washing with 600 µl wash buffer 2 per tube (Kalhor et al., 2011).
- Resuspend the beads in 100 µl wash buffer 2 per tube and keep on ice.
- Prepare a master mix for marking the DNA ends of the 4 ‘TCC’ samples by adding in the following order (Kalhor et al., 2011):
295 µl water
4.5 µl 1 M MgCl2
45 µl 10x NEBuffer 2
3.2 µl 10 mM dATP
3.2 µl 10 mM dTTP
3.2 µl 10 mM dGTPαS
68 µl 0.4 mM biotin-14-dCTP
18 µl 10% Triton X-100
11.3 µl Klenow DNA polymerase, large fragment (10 U/µl)
Mix the components gently (see Note 6) - Four ‘TCC’ samples: Add 100 µl of the master mix to the beads resuspended in 100 µl wash buffer 2.
‘3C’ control: Add 100 µl 1x NEBuffer 2 buffer (no fill-in reaction and no marking of DNA ends). - If no 3C control for biotin incorporation is desired, use the remainder 3C material for an additional, fifth TCC aliquot. Adjust the volumes of master mixes accordingly.
- Place the five samples into a tube rotator for 40 min.
- Stop the fill-in reaction by adding 5 µl 0.5 M EDTA, pH 8.0 to each tube (Kalhor et al., 2011) and mix well by inversion.
- Place the five tubes into an MPC, wait for 2 min and discard the supernatant.
- Remove the tubes from the MPC, add 600 µl wash buffer 3 (Kalhor et al., 2011) per tube and resuspend the beads by gentle vortexing.
- Place the tubes in the MPC for 2 min and discard the supernatant.
- Repeat the washing with 600 µl wash buffer 3 per tube.
- Resuspend the beads in 500 µl wash buffer 3 per tube and transfer the beads to five 15 ml conical tubes (label four tubes ‘TCC’ and one tube ‘3C’).
- Prepare a master mix by adding (Kalhor et al., 2011):
21.1 ml water
1,350 µl 10x ligation buffer TCC
972 µl 10% Triton X-100
540 µl 1 M Tris-HCl, pH 7.4
270 µl BSA (10 mg/ml) - Add 4,480 µl of the master mix to the five conical tubes (total volume 4,980 µl/ tube).
- Add 12 µl T4 DNA ligase (5 U/µl) to each of the four TCC tubes and 3 µl T4 DNA ligase (5 U/µl) to the ‘3C’ control.
- Mix the components gently by inversion.
- Incubate horizontally in a cabinet (16 °C, overnight) with agitation fast enough to keep the beads in suspension.
- Stop the ligation by adding 200 µl 0.5 M EDTA, pH 8.0 to each tube (Kalhor et al., 2011) and mix gently by inversion.
- Reclaim the beads by mounting the five 15 ml tubes horizontally on a DynaMag-96 Side Skirted Magnetic Particle Concentrator (MPC96). Fasten the tubes securely to the magnet by using tape or a rubber strap (Figure 9).

Figure 9. Recovery of tethered circularization products. Tubes containing the ligation products are mounted on a DynaMag-96 Side Skirted Magnetic Particle Concentrator. The beads are reclaimed, and the supernatant is discarded while the tubes are placed in the magnet. - Incubate for 5 min to reclaim the beads.
- Discard the supernatant using a 10 ml pipette.
- Reversion of crosslink and DNA extraction
- Add 400 µl extraction buffer (Kalhor et al., 2011) to each 15 ml tube and resuspend the beads.
- Transfer the beads to five 1.5 ml tubes (four labeled ‘TCC’ and one ‘3C’).
- Digest remaining RNA by adding 4 µl RNase A (25 µg/µl) to each 1.5 ml tube following incubation at 37 °C for 45 min.
- Add 20 µl Proteinase K (20 µg/µl) per tube, mix gently and incubate in a water bath (65 °C, overnight) (Kalhor et al., 2011).
- Add additional 5 µl Proteinase K (20 µg/µl) per tube, mix gently and incubate in a water bath (65 °C, 2 h).
- Place the five tubes into an MPC, wait for 2 min and transfer the supernatant containing the DNA into five fresh 1.5 ml tubes.
- Extract the DNA by adding 400 µl phenol:CHCl3 to each tube (Kalhor et al., 2011). Shake manually for 1 min. Separate the phases by centrifugation (room temperature, 13,000 x g, 10 min). Transfer the upper aqueous phase into a new 1.5 ml tube (see Note 7).
- Repeat the extraction with 400 µl phenol:CHCl3.
- Repeat the extraction once with 400 µl CHCl3.
- Transfer the upper aqueous phase into a new 2.0 ml tube and determine the volumes.
- For DNA precipitation add 1/20 volume 4 M NaCl, 1/100 volume glycogen (5 mg/ml) and 2 volumes ice-cold 100% ethanol to each tube (Kalhor et al., 2011) (see Note 8).
- Mix the five tubes well by inversion and incubate for 30 min on ice.
- Collect the DNA by centrifugation (4 °C, 13,000 x g, 30 min).
- Carefully remove the supernatant without disturbing the translucent DNA pellet.
- Add 500 µl ice-cold 80% ethanol and spin the DNA (4 °C, 13,000 x g, 5 min).
- Discard the supernatant. Perform a pulse-spin and remove liquid remnants.
- Dry the pellet (5 min, room temperature).
- Dissolve the five DNA pellets in 30 µl EB per tube. Pool the four TCC samples in one 1.5 ml tube. Precipitate the DNA of the TCC and 3C control by adding 1/10 volume 3 M sodium acetate, pH 5.2 and two volumes ice-cold 100% ethanol.
- Mix well by inversion and incubate for 30 min on ice.
- Collect the DNA by centrifugation (4 °C, 13,000 x g, 30 min).
- Carefully remove the supernatant from the two tubes.
- Add 500 µl ice-cold 80% ethanol.
- Spin the DNA (4 °C, 13,000 x g, 15 min).
- Discard the supernatants. Perform a pulse-spin and remove liquid remnants.
- Dry the pellets (5 min, room temperature).
- Dissolve the TCC sample in 50 µl EB and the 3C control in 10 µl EB.
- Determine the DNA concentration using the Qubit 2.0 (or Qubit 4) fluorometer and the dsDNA BR Assay. The concentration should be 100-800 ng/µl.
- The samples can be stored at -20 °C.
- Measure the biotin incorporation (Belton et al., 2012) (see Note 9).
Set up two PCR reactions for the 3C control and the TCC sample:
240 ng DNA
15 µl 5x Q5 Hot Start buffer
1.5 µl 10 mM dNTPs
3.75 µl 10 mM primer 1 + 2 (10 µM each)
0.75 µl Q5 Hot Start High-Fidelity DNA Polymerase (2U/µl)
Add water to a final volume of 75 µl - Amplify for 1 cycle (98 °C for 40 sec, 66 °C for 30 sec, 72 °C for 30 sec) followed by 35 cycles (98 °C for 10 sec, 66 °C for 30 sec, 72 °C for 30 sec) and a final amplification (98 °C for 10 sec, 66 °C for 30 sec, 72 °C for 5 min).
- Purify the products using the ‘QIAquick PCR Purification Kit’ according to the manufacturer’s instructions. Elute the PCR product in 23 µl EB.
- Set up a control and three digestions for the ‘3C control for TCC’ and ‘TCC’ (Table 1). Incubate at 37 °C (incubator cabinet, overnight).
Table 1. Digestion setup for the verification of marking and ligation of ends
- Add 4 µl 6x loading dye and analyze the products on a standard 2% agarose gel (Green and Sambrook, 2012). Typical results are shown in Figure 10.

Figure 10. Typical control for marking and ligation of ends. Two close genomic barley HindIII fragments are PCR-amplified across their ligation junction. HindIII-digestion of the amplified 3C junctions, in which HindIII sticky-ends are ligated, yields two fragments of a different size. In contrast, TCC junctions are created from the blunt-end ligation of filled-in and marked HindIII ends. As a result, novel NheI sites are formed and the original HindIII sites are lost (Belton et al., 2012). Typically, approximately 50-80% of the PCR-products are cleaved by NheI. The size of the products is indicated (bp). The gel images are taken from Mascher et al. (2017).
- Removal of biotin from non-ligated DNA ends and DNA fragmentation
- To 5 µg DNA (Step I28) add 9 µl 10x NEBuffer 1 and water to 84 µl.
- Add 6 µl Exonuclease III (100 U/µl).
- Mix gently and incubate (41 °C, 1 h) (see Note 10).
- Stop the reaction by adding 2 µl 0.5 M EDTA, pH 8.0 and 2.5 µl 4 M NaCl (Kalhor et al., 2011).
- Incubate (70 °C, 20 min) and adjust the volume with water to 100 µl.
- Shear the DNA to 100-500 bp using a Covaris S220 AFA Ultrasonicator (Duty Factor: 10%, PIP: 175 W, Cycles per burst: 200, Set Mode: Frequency sweeping, Time: 60 sec) and Covaris microTUBES.
- Control the DNA size by running 100 ng on a standard 2% agarose gel (Green and Sambrook, 2012). Add another sonication cycle, if the DNA is still too large.
- Transfer the sample completely into a 1.5 ml tube. Determine the volume exactly.
- Add 1.8 volumes AMPure XP beads (room temperature), mix, incubate (5 min).
- Reclaim the beads using a MPC (2 min).
- Wash beads for 30 sec with 190 µl 70% ethanol while the tube is placed in the MPC. Discard the supernatant.
- Repeat the wash step (Step J11) once.
- Air dry beads completely.
- Remove the tube from the MPC and add 52 µl EBT for elution.
- Resuspend the beads, incubate (1 min), place the tube in an MPC (1 min).
- Transfer 51 µl to a new tube.
- The samples can be stored at -20 °C.
- End repair and A-tailing
- Set a heat block to 20 °C.
- For end repair add to the fragmented, cleaned-up DNA in the following order:
7 µl 10x Tango
7 µl 2.5 mM dNTP mix
0.7 µl 100 mM ATP
1.5 µl T4 DNA polymerase (5 U/µl)
2.5 µl T4 polynucleotide kinase (10 U/µl)
0.3 µl Klenow DNA polymerase, large fragment (10 U/µl)
The total volume is 70 µl - Mix carefully by pipetting up and down.
- Incubate (20°C, 30 min) the sample.
- Determine the volume exactly.
- Add 1.8 volumes AMPure XP beads (room temperature), mix, incubate (5 min).
- Reclaim the beads using an MPC (2 min).
- Wash beads for 30 sec with 190 µl 70% ethanol while the tube is placed in the MPC. Discard the supernatant.
- Repeat wash step (Step K8) once.
- Air dry beads completely.
- Remove the tube from the MPC and add 39 µl TLE for elution.
- Resuspend the beads, incubate (1 min), place the tube in an MPC (1 min).
- Transfer 38 µl to a new tube.
- Add an ‘A’ by the addition of:
5 µl 10x NEBuffer 2
4 µl 2.5 mM dATP
3 µl Klenow fragment, 3’→5’ exo- (5 U/µl)
The total volume is 50 µl (Kalhor et al., 2011) - Mix carefully by pipetting up and down and incubate (37 °C, 30 min) the sample.
- During the incubation set a heat block to 65 °C.
- Inactivate the enzyme by adding 1 µl 0.5 M EDTA, pH 8.0 and incubation at 65 °C for 20 min. Place the samples on ice immediately.
- During the inactivation prepare the Dynabeads MyOne Streptavidin C1.
- Biotin pull-down
- All subsequent steps are performed in 1.5 ml DNA LoBind tubes.
- Vortex Dynabeads MyOne Streptavidin C1 and transfer 10 µl to a 1.5 ml tube.
- Add 400 µl TWB, mix by pipetting and incubate the beads on a rocking platform for 3 min.
- Reclaim the beads using an MPC (1 min) and discard the supernatant.
- Remove the tube from the MPC, add 400 µl TWB and mix by pipetting.
- Reclaim the beads using an MPC (1 min) and discard the supernatant.
- Resuspend beads in 50 µl 2x BB and add 50 µl of DNA for the affinity purification of biotinylated DNA fragments (Kalhor et al., 2011).
- Incubate for 30 min with rotation.
- Reclaim the beads using an MPC (1 min) and discard the supernatant.
- Resuspend beads in 800 µl 1x BB and transfer the suspension into a new tube.
- Reclaim the beads using an MPC (1 min) and discard the supernatant.
- Repeat washing step with 1x BB.
- Remove the tube from the MPC, add 100 µl 1x ligation buffer and mix by pipetting.
- Reclaim the beads using an MPC (1 min) and discard the supernatant.
- Remove the tube from the MPC and resuspend the beads in 38.8 µl 1x ligation buffer.
- Illumina paired-end adapter ligation and PCR
- Set up the ligation by adding to 1.5 ml tube:
6 µl TruSeq DNA Single Indexes (Set A)
38.8 µl beads with affinity purified DNA
1.1 µl 10x T4 DNA ligation buffer
1.1 µl PEG 4000 (50%)
1.0 µl water
2.0 µl T4 DNA ligase (5 U/µl).
Note: For the selection of ‘TruSeq DNA Indexes’, follow the instructions provided in the ‘TruSeq Library Pooling Guide’ (Illumina, San Diego, California, U.S.A.).
Mix well by pipetting and incubate for 1 h (22°). - Reclaim the beads using an MPC (1 min) and discard the supernatant.
- Resuspend beads in 400 µl TWB and rotate 5 min.
- Reclaim the beads using an MPC (1 min) and discard the supernatant.
- Repeat washing with 400 µl TWB twice.
- Resuspend beads in 200 µl 1x BB.
- Reclaim the beads using an MPC (1 min) and discard the supernatant.
- Resuspend beads in 200 µl 1x NEBuffer 2.
- Reclaim the beads using an MPC (1 min) and discard the supernatant.
- Repeat washing with 200 µl 1x NEBuffer 2.
- Remove the tube from the MPC.
- Resuspend the beads in 20 µl 1x NEBuffer 2.
- Transfer the sample to a new 1.5 ml tube and store the bead-bound TCC DNA on ice.
- Titrate the optimal number of PCR-cycles (Belton et al., 2012).
Resuspend the beads and add to a PCR tube:
1.5 µl bead-bound TCC DNA
1.3 µl 10 µM primer 3 + 4 (see Note 11)
5.0 µl 5x Q5 Hot Start buffer
2.0 µl 2.5 mM dNTP mix
0.3 µl Q5 Hot Start High-Fidelity DNA Polymerase (2 U/µl)
14.9 µl water (final volume 25 µl). - Titration PCR: After an initial incubation at 98 °C (30 sec), perform the amplification of the DNA (20 cycles: 98 °C for 10 sec, 68 °C for 30 sec, and 72 °C for 30 sec), followed by a final 5 min extension at 72 °C.
- Take 4 µl aliquots after 9, 12, 15 and 20 cycles.
- Analyse the aliquots using a standard agarose gel (Figure 11). Estimate the amount of products by comparing to a known DNA quantity from the ladder. Choose the number PCR cycles and numbers of reactions (25 µl volume) to produce approximately 200-600 ng DNA for sequencing (see Note 12). In order to avoid PCR artifacts indicated by changes in size distribution of the PCR products, it is recommended to use at most 15 cycles (Belton et al., 2012). The library shown in Figure 11 yielded sufficient amounts of DNA at 12 cycles.

Figure 11. Titration PCR. After the indicated number of PCR cycles, 2.4 µl of the reaction (initial volume of PCR: 25 µl) is withdrawn, size-separated using a standard 2% agarose gel and stained with ethidium bromide. The size of the GeneRuler 50 bp DNA ladder (L) is indicated (bp). The 500 bp marker band corresponds to 20 ng DNA. - Resuspend the bead-bound TCC DNA and set up the final PCR with the calculated number of reactions. Perform PCR using the optimal number of cycles.
- Set up the ligation by adding to 1.5 ml tube:
- Reaction clean-up and size selection
- Pool all PCR-products in one 1.5 ml tube and place the tube in an MPC.
- Wait for 1 min and transfer the supernatant containing the library into a new 1.5 ml tube for clean-up and size selection (Belton et al., 2012).
- Determine the volume exactly.
- Add 1.8 volumes AMPure XP beads (room temperature), mix, incubate (5 min) and reclaim the beads using an MPC (2 min).
- Wash beads for 30 sec with 190 µl 70% ethanol while the tube is placed in the MPC.
- Discard the supernatant.
- Repeat the wash step (Steps N5-N6) once.
- Air dry beads completely.
- Remove the tube from the MPC and add 27 µl EBT for elution.
- Resuspend the beads and incubate (10 min). Tap the tube every 2 min to resuspend the beads.
- Place the tube in an MPC (1 min) and transfer 25 µl to a new tube.
- Determine the DNA concentration using the Qubit 2.0 (or Qubit 4) fluorometer and the dsDNA BR Assay. The concentration should be 10-30 ng/µl.
- Use a gel-purification to make sure that appropriately sized DNA fragments are used for sequencing (Himmelbach et al., 2014) (see Note 13). Weigh 1 g UltraPure Agarose in a 500 ml Erlenmeyer flask. Add 50 ml 1x TAE buffer and melt the agarose using a microwave until dissolved completely. Cool the liquid to 60 °C, add 5 µl SYBR-Gold (see Note 14), mix gently without forming bubbles and cast the gel using standard electrophoresis equipment. Cover the gel with aluminum foil and let the gel solidify (light-sensitive dye).
- Transfer 250 ng GeneRuler 50 bp DNA ladder into a tube, add 4 µl 6x loading dye and adjust the volume to 24 µl using EBT. Prepare the TCC sample (25 µl) by adding 5 µl 6x loading dye.
- Remove the comb and place the gel in an electrophoresis chamber filled with 1x TAE buffer. Load the TCC library in two adjacent slots. Apply the standard to one lane and perform electrophoresis for 1 h with 5 V/cm (distance between anode and cathode). Protect the gel from light during separation. Visualize the DNA using a ‘Dark reader’ transilluminator (see Note 15) and take a photograph. Use a clean scalpel and excise the region between 300 and 550 bp (Figure 12).

Figure 12. Gel-purification of a typical TCC library. Following size-separation (standard 2% agarose gel electrophoresis) and staining with SYBR Gold, the library is revealed using a visible blue light emitting ‘Dark reader transilluminator’. The DNA from the framed area between approximately 300 and 550 bp is extracted for sequencing. The size of the GeneRuler 50 bp DNA ladder (L) is indicated (bp). - Place the gel block in a 2.0 ml tube and determine the volume using a balance (e.g., block of 234 mg corresponds to 234 µl).
- Purify the DNA using the ‘MinElute Gel Extraction Kit’ essentially as described by the manufacturer. Add 6 volumes QG buffer and dissolve the gel completely under gentle agitation.
- Add one gel volume isopropanol and mix by inversion.
- Apply 700 μl of the dissolved gel to a MinElute column. Place the column in a collection tube and spin (16,000 x g, 1 min). Discard the flow-through.
- Stepwise add the residual liquid to the column (repeat Step N19).
- Add 740 μl PE to wash the column, incubate for 3 min, spin the column (16,000 x g, 1 min) and discard the flow-through (see Note 16).
- Turn the column (180°) in the rotor and spin (16,000 x g, 1 min).
- Discard the collection tube, place the MinElute column into a clean 1.5 ml tube and for elution add 50 μl EB to the resin.
- Incubate for 1 min and spin the column (16,000 x g, 1 min).
- Store the eluted TCC library at -20 °C.
- Quality controls, quantification and sequencing
- Measure the DNA concentration using the Qubit 2.0 (or Qubit 4) fluorometer (ds DNA HS Assay). The concentration should be 10-20 ng/µl.
- Digest 80 ng TCC library with NheI to estimate the fraction derived from ligation of biotinylated junctions (Belton et al., 2012). Set up the reaction by adding to a 1.5 ml tube:
80 ng TCC library
2 µl 10x Tango buffer
1 µl NheI (10 U/µl)
Water to a final volume of 20 µl
Prepare an uncut control (without NheI digestion). Incubate overnight at 37 °C. Analyse the products with a 2% agarose gel using 2 µl GeneRuler 50 bp DNA ladder as a standard. The smaller size distribution of the digested TCC library provides an estimate for the fraction containing true TCC ligation products (Figure 13).
Figure 13. Quality control of a final TCC library. After digestion with NheI the TCC library (80 ng) is size-separated (standard 2% agarose gel), stained with ethidium bromide and compared to the uncut control (80 ng). The shift to a smaller size distribution is indicative of the presence of NheI sites, which originate from genuine TCC ligation events (Belton et al., 2012; Mascher et al., 2017). In general, we found that the shift is a secure quality indicator. Libraries without a clear shift should not be sequenced, because they will yield only a very small fraction of useful TCC reads. The size of the GeneRuler 50 bp DNA ladder (L) is indicated (bp). - Record the size profile of the library and determine the average size of the TCC library using the Agilent High Sensitivity DNA Kit with the Agilent 2100 Bioanalyzer (Figure 14).

Figure 14. Size profile of a typical TCC library. The TCC library is size-fractionated using the Agilent 2100 Bioanalyzer. The average library size (Region 1: 434 bp; including 121 bp Illumina P5 and P7 adapter) corresponds to an average insert size of about 313 bp. LM: lower marker peak. UM: upper marker peak. FU: fluorescence unit. bp: base pairs. - Quantify and sequence the library (paired-end, 2x 100 cycles) using an Illumina system as described (Mascher et al., 2013).
Data analysis
Primary sequence data analysis
- Run the Illumina CASAVA pipeline (http://support.illumina.com/sequencing/sequencing_software/casava.html) to obtain deconvoluted read files in FASTQ format.
- Trim the reads at the junction site with cutadapt (Martin, 2011) using the adapter sequence 'AAGCTAGCTT'.
- Align the trimmed read pairs to an appropriate reference genome with BWA mem (Li, 2013). The two reads should be mapped as single ends by specifying the parameters ‘-S’ and ‘-P’. The parameter ‘-M’ should be used to mark shorter split hits as secondary.
- Sort BAM files by reference position for duplicates removal. Then sort the BAM files again by read name to have the two ends of a pair at adjacent rows of the BAM file. One of the tools SAMtools (Li et al., 2009), Picard (http://broadinstitute.github.io/picard/) or Novosort (http://www.novocraft.com/products/novosort/) can be used.
- Remove reads with non-unique alignments and discard secondary alignments, unmapped reads and duplicated reads with command 'samtools view -q 10 -F1284'.
- Assign reads to restriction fragments with BEDTools (Quinlan and Hall, 2010). You will need a BED file (https://genome.ucsc.edu/FAQ/FAQformat.html#format1) with the positions of restriction fragments (i.e., regions between two HindIII sites) obtained from a HindIII in silico digest of your reference genome sequence. Use the command ‘bedtools pairtobed -bedpe -f 1 -type both -abam’. The resulting BEDPE file is a tab-separated file that can be processed with standard UNIX text processing tools such as AWK or Perl. The assignments of the two ends of a pair to restriction fragments are on adjacent lines.
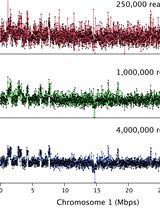
- Use UNIX scripts (e.g., in the AWK language) to calculate the insert size based on the distance of alignment start positions to neighboring HindIII sites. Discard fragments with insert sizes above 500 bp, plot the insert size distribution (Figure 15) and compare it to that obtained with the Agilent Bioanalyzer.

Figure 15. Typical in silico size profile of a TCC library. The sequence data are mapped to a reference genome and assigned to restriction fragments. The insert size of each sequenced fragments is determined from the distance of alignment start positions to the next HindIII restriction site. The mode of the distribution is indicated by a red line.
Notes
- Approach the local sales representative for ordering, performance and accessory items.
- If darkening of the leaves is not observed, the fixative might not have entered completely, thus indicating an incomplete crosslink. Optimize the infiltration conditions for example by increasing the vacuum or cutting shorter plant segments.
- It is mandatory to wear appropriate protective gear. Consult the Safety Officer of your institution for proper handling of liquid nitrogen.
- The MyOne Streptavidin T1 beads provide about 250 cm2 surface per ml. By using 400 µl beads, the purified chromatin is tethered at low density on a large surface of about 100 cm2 (Kalhor et al., 2011). Thereby intramolecular ligation of DNA ends is favored.
- Free streptavidin residues are saturated with biotin in order to avoid interference with biotin-14-dCTP, which is used for marking the DNA ends during the following step (Kalhor et al., 2011).
- During this reaction blunt DNA ends are generated, which are labeled with a biotinylated cytosine residue located 3’ to a phosphorothioate bond introduced by the incorporation of dGTPαS (Kalhor et al., 2011).
- Spin the samples in a centrifuge equilibrated to room temperature. Cold temperatures might result in the formation of a turbid water phase. Avoid contaminating the water phase with protein from the interphase during the transfer.
- NaCl is used for DNA precipitation, because the solution contains SDS (Green and Sambrook, 2012). Subsequently, the DNA is precipitated in the presence of sodium acetate to remove traces of sodium chloride, which is inhibitory to Exonuclease III (Hoheisel, 1993).
- Ligation of filled-in HindIII sites (AAGCTT) generates sites for the restriction enzyme NheI (GCTAGC). To control for effectiveness of fill-in and blunt-end ligation, a PCR fragment (534 bp) from two adjacent barley HindIII restriction fragments is generated. For amplification, primer 1 and primer 2 are used.
- Exonuclease III from E. coli catalyzes the stepwise removal of mononucleotides from 3’-hydroxyl termini of duplex DNA (New England BioLabs; Rogers and Weiss, 1980). Thus, Exonuclease III allows for the removal of biotinylated residues from the non-ligated DNA ends. Because phosphorothioate linkages located 5’ to the biotinylated cytosine residue are not cleaved by Exonuclease III (Putney et al., 1981), true ligation junctions are preserved (Kalhor et al., 2011).
- Primers 3 and 4 anneal to the Illumina adapter and allow for the amplification of the library.
- In routine experiments 11-12 amplification cycles are sufficient. Eight PCR reactions (25 µl volume per reaction) should yield enough DNA for sequencing. Follow the ‘golden rule’ to sustain the complexity of libraries: Keep the number of cycles low and increase the number of PCR reactions to obtain sufficient library for sequencing (Belton et al., 2012).
- The BluePippin from Sage Science is an alternative to standard gel-purifications.
- Do not use ethidium bromide stained gels. Ultra-violet radiation used for excitation of ethidium bromide will damage the DNA.
- DNA is revealed in agarose gels using SYBR-Gold dye and visible blue light emitted from a 'Dark reader' transilluminator as the excitation source.
- Place the columns in the rotor consistently, since the columns will be turned 180° for removing traces of liquid.
Recipes
Notes:
- All solutions are prepared using purified water (GenPure Pro UV/UF) and stored at room temperature unless specified otherwise.
- Good laboratory practice has to be applied throughout the experiment.
- It is especially important to consult the Safety Data Sheets and the laboratory/environmental safety rules of your institution for proper handling the reagents and instruments.
- 1 M HEPES, pH 8.0 (100 ml)
- Weigh 26.0 g HEPES sodium salt (4-(2-Hydroxyethyl) piperazine-1-ethanesulfonic acid sodium salt) and add 70 ml water
- Stir and adjust the pH to 8.0 with concentrated HCl using a pH meter. Adjust to 100 ml with water
- Sterilize using a 0.2 µm filter and a 50 ml syringe (store solution in darkness at room temperature)
- 2 M sucrose (40 ml)
- Fill 27.4 g sucrose into a 50 ml tube
- Add water to a volume of 38 ml and place the 50 ml tube in a warm water bath
- Vortex the warm mixture in regular intervals to dissolve the sucrose completely
- Adjust the volume to 40 ml and store the solution at -20 °C
- Thaw in a warm water bath prior to use
- 1 M KCl (100 ml)
- Dissolve 7.5 g KCl in water to a final volume of 100 ml
- Use a graduated cylinder to adjust the volume and sterilize by autoclaving
- Store at room temperature
- 10% (v/v) Triton X-100 (45 ml)
- Add 5 ml Triton X-100 to 45 ml water into a 50 ml tube
- Mix gently to avoid foaming
- Store 10% Triton X-100 and solutions containing Triton X-100 protected from light at room temperature
- Nuclei Isolation Buffer (NIBF) with formaldehyde (Hövel et al., 2012) (200 ml)
Add all of the components into a 250 ml bottle and store on ice before use:
4 ml 1 M HEPES, pH 8.0
25 ml 2 M sucrose
200 µl 1 M MgCl2
1 ml 1 M KCl
100.8 g 100% (v/v) glycerol (corresponding to 80 ml)
5 ml 10% (v/v) Triton X-100
10.7 ml 37% (w/v) formaldehyde
200 µl 100 mM PMSF
0.1% (v/v) 2-mercaptoethanol
Add water to a final volume of 200 ml
Note: PMSF inhibits serine proteases and is unstable in water. Due to the short half-life of 35 min at pH 8.0 (James, 1978), PMSF must be added immediately before use. In the air formaldehyde slowly oxidizes to formic acid. Buy small quantities and use them shortly after purchase. Poor quality formaldehyde will spoil the experiment. Store formaldehyde at room temperature, because in the cold a precipitate may be formed. Formaldehyde, PMSF and 2-mercaptoethanol are added inside the fume hood just before use. Stir thoroughly and chill the solution on ice. Use NIBF immediately after composition for vacuum infiltration. 100% glycerol (density 1.26 g/ml) is viscous and sticks to pipettes and measuring cylinders. For convenience, weigh 100.8 g 100% glycerol corresponding to 80 ml in the glass bottle. - 2 M glycine (40 ml)
- Weigh 6.0 g glycine into a 50 ml tube and adjust to 40 ml with water
- Store at -20 °C and thaw in a warm water bath prior to use
- 1 mg/ml Aprotinin (Haring et al., 2007)
- Dissolve 1 mg Apronitin in 1 ml water
- Store in aliquots at -20 °C
- 1 mg/ml Leupeptin (Haring et al., 2007)
- Dissolve 1 mg Leupeptin in 1 ml water
- Store in aliquots at -20 °C
- 1 mg/ml Pepstatin (Haring et al., 2007)
- Dissolve 1 mg Pepstatin in 1 ml water
- Store in aliquots at -20 °C
- Nuclei Isolation Buffer with protease inhibitors (NIBP) (Hövel et al., 2012) (150 ml)
Add all of the components into a 250 ml bottle and store on ice before use:
3 ml 1 M HEPES, pH 8.0
18.8 ml 2 M sucrose
150 µl 1 M MgCl2
750 µl 1 M KCl
75.6 g 100% (v/v) glycerol (corresponding to 60 ml)
3.8 ml 10% (v/v) Triton X-100
150 µl 100 mM PMSF
150 µl 100% (v/v) 2-mercaptoethanol
150 µl 1 mg/ml Aprotinin
150 µl 1 mg/ml Leupeptin
150 µl 1 mg/ml Pepstatin
Add water to a final volume of 150 ml
Note: PMSF and 2-mercaptoethanol are added inside the fume hood just before use. Stir thoroughly and chill the solution on ice. Use NIBP immediately after composition. 100% glycerol (density 1.26 g/ml) is viscous and sticks to pipettes and measuring cylinders. For convenience, weigh 75.6 g 100% glycerol corresponding to 60 ml in the glass bottle. - Sucrose cushion (20 ml)
Add all of the components into a 50 ml plastic tube and store on ice before use:
400 µl 1 M HEPES, pH 8.0
16.9 ml 2 M sucrose
20 µl 1 M MgCl2
100 µl 1 M KCl
500 µl 10% (v/v) Triton X-100
20 µl 100 mM PMSF
20 µl 100% (v/v) 2-mercaptoethanol
20 µl 1 mg/ml Aprotinin
20 µl 1 mg/ml Leupeptin
20 µl 1 mg/ml Pepstatin
Add water to a final volume of 20 ml
Note: PMSF and 2-mercaptoethanol are added inside the fume hood just before use. Stir thoroughly and chill the solution on ice. Use the sucrose cushion immediately after composition. - Nuclei Isolation Buffer with 1.5 M sucrose (NIBS) (20 ml)
Add all of the components into a 50 ml plastic tube and store on ice before use:
400 µl 1 M HEPES, pH 8.0
14.9 ml 2 M sucrose
20 µl 1 M MgCl2
100 µl 1 M KCl
500 µl 10% (v/v) Triton X-100
20 µl 100 mM PMSF
20 µl 100% (v/v) 2-mercaptoethanol
20 µl 1 mg/ml Aprotinin
20 µl 1 mg/ml Leupeptin
20 µl 1 mg/ml Pepstatin
Add water to a final volume of 20 ml
Note: PMSF and 2-mercaptoethanol are added inside the fume hood just before use. Stir thoroughly and chill the solution on ice. Use NIBS immediately after composition. - 2% (w/v) SDS (100 ml)
- Dissolve 2 g SDS (Sodium Dodecyl Sulfate) in 70 ml water
- Mix gently, avoid foaming and use a graduated cylinder to adjust the volume to 100 ml
- The solution can be stored at room temperature
- 4 M NaCl (100 ml)
- Dissolve 23.34 g NaCl in 80 ml water
- Use a graduated cylinder to adjust the volume to 100 ml, sterilize by autoclaving and store at room temperature
- Wash buffer 1 (50 ml)
Add all of the components into a 50 ml plastic tube:
2.5 ml 1 M Tris-HCl, pH 8.0
625 µl 4 M NaCl
100 µl 0.5 M EDTA, pH 8.0
Adjust with water to a final volume of 50 ml (Kalhor et al., 2011) - 25 mM IPB (150 µl)
Dissolve 2 mg EZlink Iodoacetyl-PEG2-Biotin (IPB) in wash buffer 1 (final volume 150 µl)
Note: Prepare the solution freshly before use. IPB is moisture and light sensitive. Store the reagent at 4-8 °C desiccated in a closed dark container together with dry silicate beads. Prior to opening, equilibrate the vial to room temperature to prevent the condensation of moisture in the vial. - 10x NEBuffer 2 (New England BioLabs) (10 ml)
Add all of the components:
1.25 ml 4 M NaCl
1 ml 1 M Tris-HCl, pH 8.0
1 ml 1 M MgCl2
100 µl 1 M DTT
Add water to 10 ml and store in aliquots at -20 °C - 1x NEBuffer 2
Prepare by diluting 1 ml 10x NEBuffer 2 stock with 9 ml water
Store aliquots at -20 °C - HindIII (10 U/µl)
Add all of the components on ice and mix gently:
0.5 µl HindIII (100 U/µl)
4.5 µl 1x NEBuffer 2
Prepare freshly before use - Dialysis buffer (TE buffer) (2 L)
Add all of the components:
20 ml 1 M Tris-HCl, pH 8.0
4 ml 0.5 M EDTA, pH 8.0
Add water to final volume of 2 L - Phosphate Buffered Saline with Tween 20, pH 7.4 (PBST) (1 L)
Add all of the components:
8.0 g NaCl
0.2 g KCl
1.44 g Na2HPO4
0.24 g KH2PO4
Add water to 800 ml
Adjust the pH to 7.4 using HCl (Green and Sambrook, 2012)
Add 1 ml 10% Tween 20 and water to a final volume of 1 L - 25 mM neutralized IPB (75 µl)
- Dissolve 1 mg EZlink Iodoacetyl-PEG2-Biotin (IPB) in wash buffer 1 (final volume 75 µl)
- Take 40 µl 25 mM IPB and add 7 µl 143 mM 2-mercaptoethanol
- 10 mM dATP
Dilute 100 µl 100 mM dATP with 900 µl water and store at -20 °C - 10 mM dTTP
Dilute 100 µl 100 mM dTTP with 900 µl water and store at -20 °C - Wash buffer 2 (50 ml) (Kalhor et al., 2011)
Add all of the components into a 50 ml plastic tube:
2.5 ml 1 M Tris-HCl, pH 8.0
625 µl 4 M NaCl
2 ml 10% (v/v) Triton X-100
Adjust with water to a final volume of 50 ml - Wash buffer 3 (50 ml) (Kalhor et al., 2011)
Add all of the components into a 50 ml plastic tube:
2.5 ml 1 M Tris-HCl, pH 7.4
10 µl 0.5 M EDTA, pH 8.0
2 ml 10% (v/v) Triton X-100
Adjust with water to a final volume of 50 ml - 10x ligation buffer TCC (1.5 ml)
Add all of the components into a 2.0 ml plastic tube:
750 µl 1 M Tris-HCl, pH 7.5
150 µl 1 M MgCl2
150 µl 100 mM ATP
150 µl 1 M DTT
300 µl water
Notes:- Prepare the solution freshly before use.
- The composition is identical to the 10x reaction buffer from New England BioLabs.
- 10 mg/ml BSA
- Transfer 200 µl Ultrapure BSA (50 mg/ml) into a tube
- Add 800 µl water, mix and store at -20 °C
- Extraction buffer (50 ml) (Kalhor et al., 2011)
Add all of the components:
2.5 ml 1 M Tris-HCl, pH 8.0
100 µl 0.5 M EDTA, pH 8.0
1.25 ml 4 M NaCl
1 ml 10% (v/v) SDS
Adjust with water to a final volume of 50 ml - 25 mg/ml RNase A (1 ml)
Dissolve 25 mg RNase A (DNase-free) in water in 1 ml final volume and store at 4 °C - 20 mg/ml Proteinase K (150 µl)
Dissolve 3 mg Proteinase K in 150 µl 10 mM Tris-HCl, pH 8.0
Prepare solution freshly just before use - 5 mg/ml glycogen (100 µl)
Dilute 25 µl glycogen (20 mg/ml) with 75 µl water and store at -20 °C - 80% ethanol (100 ml)
Mix 80 ml 100% ethanol and 20 ml water
Prepare freshly and store in a tightly closed bottle
Pre-cool on ice before use. - 70% ethanol (100 ml)
Mix 70 ml 100% ethanol and 30 ml water
Prepare freshly and store in a tightly closed bottle
Pre-cool on ice before use. - 3 M sodium acetate, pH 5.2 (100 ml)
Prepare 100 ml solution as described in Green and Sambrook (2012). - EB (50 ml)
Add all of the components:
500 µl 1 M Tris-HCl, pH 8.0
49.5 ml water - EBT (50 ml)
Add all of the components:
500 µl 1 M Tris-HCl, pH 8.0
250 µl 10% Tween 20
49.25 ml water - Primer 1 + 2 (10 µM each) (100 µl)
Primer 1 (100 µM, 5’- ATCTTCATGCGAGGCAGAGT -3’)
Primer 2 (100 µM, 5’- ACCGTTGAACCATCTTCAGG -3’)
Add all of the components:
10 µl 100 µM primer 1 (100 µM)
10 µl 100 µM primer 2 (100 µM)
80 µl water
Store the primer mix (biotin incorporation control) for barley (Mascher et al., 2017) at -20 °C - 10 mM dNTP mix (1 ml)
Add 400 µl dNTP mix (25 mM each) to 600 µl water and store at -20 °C - 2.5 mM dNTP mix (1 ml)
Add 100 µl dNTP mix (25 mM each) to 900 µl water and store at -20 °C - TLE (50 ml)
- Tween Wash Buffer (TWB) (50 ml) (Lieberman-Aiden et al., 2009)
Add all of the components:
250 µl 1 M Tris-HCl, pH 8.0
50 µl 0.5 M EDTA, pH 8.0
250 µl 10% Tween 20
12.5 ml 4 M NaCl
Adjust with water to a final volume of 50 ml - 2x Binding Buffer (2x BB) (50 ml) (Lieberman-Aiden et al., 2009)
Add all of the components:
500 µl 1 M Tris-HCl, pH 8.0
100 µl 0.5 M EDTA, pH 8.0
25 ml 4 M NaCl
Adjust with water to a final volume of 50 ml - 1x Binding Buffer (1x BB) (20 ml) (Lieberman-Aiden et al., 2009)
Dilute 10 ml 2x BB with 10 ml water - 1x ligation buffer (1 ml)
Add all of the components:
90 µl 10x T4 DNA Ligation buffer
90 µl 50% PEG 4000
820 µl water
Store at -20 °C - Primer 3 + 4 (10 µM each) (100 µl)
Primer 3 (100 µM, 5’- AATGATACGGCGACCACCGAGAT-3’)
Primer 4 (100 µM, 5’- CAAGCAGAAGACGGCATACGA-3’)
Add all of the components:
10 µl 100 µM primer 3 (100 µM)
10 µl 100 µM primer 4 (100 µM)
80 µl water
Store the Illumina library amplification primer mix at -20 °C - 6x loading dye (1 ml)
Add all of the components:
700 µl water
300 µl 100% (v/v) glycerol
A little bit of Bromophenol Blue
Note: Add Bromophenol Blue to obtain a light blue color. Overstaining will mask DNA (area between 200-300 bp) in standard agarose gels. - 50x TAE (1 L) (Green and Sambrook, 2012)
- Dissolve 242 g Tris base in deionized water
- Add 57.1 ml glacial acetic acid and 100 ml 0.5 M EDTA, pH 8.0
- Adjust the volume to 1 L and mix thoroughly
- The 50x TAE stock is diluted with water to 1x TAE working solution and mixed thoroughly
Acknowledgments
This work was financially supported by core funding of the Leibniz Institute of Plant Genetics and Crop Plant Research (IPK), Gatersleben, and project funding (‘TRITEX’) from the German Federal Ministry of Education and Research (BMBF, FKZ 0315954) to Nils Stein. The graphic design of Karin Lipfert is gratefully acknowledged. The authors thank Lala Aliyeva-Schnorr for help with microscopic analysis of nuclei. This protocol was adapted for the most part from wet-lab procedures (Hövel et al., 2012; Lieberman-Aiden et al., 2009; Kalhor et al., 2011; Belton et al., 2012) and from bioinformatics procedures described previously (Mascher et al., 2017; Beier et al., 2017). The authors declare no competing interests or conflicts of interest.
References
- Abdalla, K. O., Thomson, J. A. and Rafudeen, M. S. (2009). Protocols for nuclei isolation and nuclear protein extraction from the resurrection plant Xerophyta viscosa for proteomic studies. Anal Biochem 384(2): 365-367.
- Avni, R., Nave, M., Barad, O., Baruch, K., Twardziok, S. O., Gundlach, H., Hale, I., Mascher, M., Spannagl, M., Wiebe, K., Jordan, K. W., Golan, G., Deek, J., Ben-Zvi, B., Ben-Zvi, G., Himmelbach, A., MacLachlan, R. P., Sharpe, A. G., Fritz, A., Ben-David, R., Budak, H., Fahima, T., Korol, A., Faris, J. D., Hernandez, A., Mikel, M. A., Levy, A. A., Steffenson, B., Maccaferri, M., Tuberosa, R., Cattivelli, L., Faccioli, P., Ceriotti, A., Kashkush, K., Pourkheirandish, M., Komatsuda, T., Eilam, T., Sela, H., Sharon, A., Ohad, N., Chamovitz, D. A., Mayer, K. F. X., Stein, N., Ronen, G., Peleg, Z., Pozniak, C. J., Akhunov, E. D. and Distelfeld, A. (2017). Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 357(6346): 93-97.
- Beier, S., Himmelbach, A., Colmsee, C., Zhang, X. Q., Barrero, R. A., Zhang, Q., Li, L., Bayer, M., Bolser, D., Taudien, S., Groth, M., Felder, M., Hastie, A., Simkova, H., Stankova, H., Vrana, J., Chan, S., Munoz-Amatriain, M., Ounit, R., Wanamaker, S., Schmutzer, T., Aliyeva-Schnorr, L., Grasso, S., Tanskanen, J., Sampath, D., Heavens, D., Cao, S., Chapman, B., Dai, F., Han, Y., Li, H., Li, X., Lin, C., McCooke, J. K., Tan, C., Wang, S., Yin, S., Zhou, G., Poland, J. A., Bellgard, M. I., Houben, A., Dolezel, J., Ayling, S., Lonardi, S., Langridge, P., Muehlbauer, G. J., Kersey, P., Clark, M. D., Caccamo, M., Schulman, A. H., Platzer, M., Close, T. J., Hansson, M., Zhang, G., Braumann, I., Li, C., Waugh, R., Scholz, U., Stein, N. and Mascher, M. (2017). Construction of a map-based reference genome sequence for barley, Hordeum vulgare L. Sci Data 4: 170044.
- Belton, J. M., McCord, R. P., Gibcus, J. H., Naumova, N., Zhan, Y. and Dekker, J. (2012). Hi-C: a comprehensive technique to capture the conformation of genomes. Methods 58(3): 268-276.
- Burton, J. N., Adey, A., Patwardhan, R. P., Qiu, R., Kitzman, J. O. and Shendure, J. (2013). Chromosome-scale scaffolding of de novo genome assemblies based on chromatin interactions. Nat Biotechnol 31(12): 1119-1125.
- Dekker, J., Rippe, K., Dekker, M. and Kleckner, N. (2002). Capturing chromosome conformation. Science 295(5558): 1306-1311.
- de Wit, E. and de Laat, W. (2012). A decade of 3C technologies: insights into nuclear organization. Genes Dev 26(1): 11-24.
- Duan, Z., Andronescu, M., Schutz, K., McIlwain, S., Kim, Y. J., Lee, C., Shendure, J., Fields, S., Blau, C. A. and Noble, W. S. (2010). A three-dimensional model of the yeast genome. Nature 465(7296): 363-367.
- Dudchenko, O., Batra, S. S., Omer, A. D., Nyquist, S. K., Hoeger, M., Durand, N. C., Shamim, M. S., Machol, I., Lander, E. S., Aiden, A. P. and Aiden, E. L. (2017). De novo assembly of the Aedes aegypti genome using Hi-C yields chromosome-length scaffolds. Science 356(6333): 92-95.
- Feng, S., Cokus, S. J., Schubert, V., Zhai, J., Pellegrini, M. and Jacobsen, S. E. (2014). Genome-wide Hi-C analyses in wild-type and mutants reveal high-resolution chromatin interactions in Arabidopsis . Mol Cell 55(5): 694-707.
- Green, M. and Sambrook, J. (2012). Molecular cloning. A laboratory manual. Cold Spring Harbor Laboratory, New York.
- Grob, S., Schmid, M. W. and Grossniklaus, U. (2014). Hi-C analysis in Arabidopsis identifies the KNOT, a structure with similarities to the flamenco locus of Drosophila. Mol Cell 55(5): 678-693.
- Haring, M., Offermann, S., Danker, T., Horst, I., Peterhansel, C. and Stam, M. (2007). Chromatin immunoprecipitation: optimization, quantitative analysis and data normalization. Plant Methods 3: 11.
- Himmelbach, A., Knauft, M. and Stein, N. (2014). Plant sequence capture optimised for illumina sequencing. Bio-protocol 4(13): e1166.
- Hoheisel, J. D. (1993). On the activities of Escherichia coli exonuclease III. Anal Biochem 209(2): 238-246.
- Hövel, I., Louwers, M. and Stam, M. (2012). 3C Technologies in plants. Methods 58(3): 204-211.
- James, G. T. (1978). Inactivation of the protease inhibitor phenylmethylsulfonyl fluoride in buffers. Anal Biochem 86(2): 574-579.
- Kalhor, R., Tjong, H., Jayathilaka, N., Alber, F. and Chen, L. (2011). Genome architectures revealed by tethered chromosome conformation capture and population-based modeling. Nat Biotechnol 30(1): 90-98.
- Kaplan, N. and Dekker, J. (2013). High-throughput genome scaffolding from in vivo DNA interaction frequency. Nat Biotechnol 31(12): 1143-1147.
- Li, H. (2013). Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv preprint arXiV: 1303.3997.
- Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., Marth, G., Abecasis, G., Durbin, R. and Genome Project Data Processing, S. (2009). The sequence alignment/map format and SAMtools. Bioinformatics 25(16): 2078-2079.
- Lieberman-Aiden, E., van Berkum, N. L., Williams, L., Imakaev, M., Ragoczy, T., Telling, A., Amit, I., Lajoie, B. R., Sabo, P. J., Dorschner, M. O., Sandstrom, R., Bernstein, B., Bender, M. A., Groudine, M., Gnirke, A., Stamatoyannopoulos, J., Mirny, L. A., Lander, E. S. and Dekker, J. (2009). Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science 326(5950): 289-293.
- Liu, C., Wang, C., Wang, G., Becker, C., Zaidem, M. and Weigel, D. (2016). Genome-wide analysis of chromatin packing in Arabidopsis thaliana at single-gene resolution. Genome Res 26(8): 1057-1068.
- Martin, M. (2011). Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal v. 17, n. 1, p. pp. 10-12, may. ISSN 2226-6089.
- Mascher, M., Richmond, T. A., Gerhardt, D. J., Himmelbach, A., Clissold, L., Sampath, D., Ayling, S., Steuernagel, B., Pfeifer, M., D'Ascenzo, M., Akhunov, E. D., Hedley, P. E., Gonzales, A. M., Morrell, P. L., Kilian, B., Blattner, F. R., Scholz, U., Mayer, K. F., Flavell, A. J., Muehlbauer, G. J., Waugh, R., Jeddeloh, J. A. and Stein, N. (2013). Barley whole exome capture: a tool for genomic research in the genus Hordeum and beyond. Plant J 76(3): 494-505.
- Mascher, M., Gundlach, H., Himmelbach, A., Beier, S., Twardziok, S. O., Wicker, T., Radchuk, V., Dockter, C., Hedley, P. E., Russell, J., Bayer, M., Ramsay, L., Liu, H., Haberer, G., Zhang, X. Q., Zhang, Q., Barrero, R. A., Li, L., Taudien, S., Groth, M., Felder, M., Hastie, A., Simkova, H., Stankova, H., Vrana, J., Chan, S., Munoz-Amatriain, M., Ounit, R., Wanamaker, S., Bolser, D., Colmsee, C., Schmutzer, T., Aliyeva-Schnorr, L., Grasso, S., Tanskanen, J., Chailyan, A., Sampath, D., Heavens, D., Clissold, L., Cao, S., Chapman, B., Dai, F., Han, Y., Li, H., Li, X., Lin, C., McCooke, J. K., Tan, C., Wang, P., Wang, S., Yin, S., Zhou, G., Poland, J. A., Bellgard, M. I., Borisjuk, L., Houben, A., Dolezel, J., Ayling, S., Lonardi, S., Kersey, P., Langridge, P., Muehlbauer, G. J., Clark, M. D., Caccamo, M., Schulman, A. H., Mayer, K. F. X., Platzer, M., Close, T. J., Scholz, U., Hansson, M., Zhang, G., Braumann, I., Spannagl, M., Li, C., Waugh, R. and Stein, N. (2017). A chromosome conformation capture ordered sequence of the barley genome. Nature 544(7651): 427-433.
- Metzker, M. L. (2010). Sequencing technologies - the next generation. Nat Rev Genet 11(1): 31-46.
- Naumova, N., Imakaev, M., Fudenberg, G., Zhan, Y., Lajoie, B. R., Mirny, L. A. and Dekker, J. (2013). Organization of the mitotic chromosome. Science 342(6161): 948-953.
- Putney, S. D., Benkovic, S. J. and Schimmel, P. R. (1981). A DNA fragment with an alpha-phosphorothioate nucleotide at one end is asymmetrically blocked from digestion by exonuclease III and can be replicated in vivo . Proc Natl Acad Sci U S A 78(12): 7350-7354.
- Quinlan, A. R. and Hall, I. M. (2010). BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26(6): 841-842.
- Rao, S. S., Huntley, M. H., Durand, N. C., Stamenova, E. K., Bochkov, I. D., Robinson, J. T., Sanborn, A. L., Machol, I., Omer, A. D., Lander, E. S. and Aiden, E. L. (2014). A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell 159(7): 1665-1680.
- Rogers, G. and Weiss, B. (1980). In: Grossman, L. and Moldave, K. (Eds.). [26] Exonuclease III of Escherichia coli K-12, an AP endonuclease. In: Methods in enzymol. 65: 201-211. New York: Academic Press.
- Sexton, T., Yaffe, E., Kenigsberg, E., Bantignies, F., Leblanc, B., Hoichman, M., Parrinello, H., Tanay, A. and Cavalli, G. (2012). Three-dimensional folding and functional organization principles of the Drosophila genome. Cell 148(3): 458-472.
- Wang, C., Liu, C., Roqueiro, D., Grimm, D., Schwab, R., Becker, C., Lanz, C. and Weigel, D. (2015). Genome-wide analysis of local chromatin packing in Arabidopsis thaliana. Genome Res 25(2): 246-256.
- Zhou, X., Lowdon, R. F., Li, D., Lawson, H. A., Madden, P. A., Costello, J. F. and Wang, T. (2013). Exploring long-range genome interactions using the WashU Epigenome Browser. Nat Methods 10(5): 375-376.
- Zimmermann, G., Baumlein, H., Mock, H. P., Himmelbach, A. and Schweizer, P. (2006). The multigene family encoding germin-like proteins of barley. Regulation and function in Basal host resistance. Plant Physiol 142(1): 181-192.
Article Information
Copyright
© 2018 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Himmelbach, A., Walde, I., Mascher, M. and Stein, N. (2018). Tethered Chromosome Conformation Capture Sequencing in Triticeae: A Valuable Tool for Genome Assembly. Bio-protocol 8(15): e2955. DOI: 10.21769/BioProtoc.2955.
Category
Plant Science > Plant molecular biology > DNA > DNA sequencing
Molecular Biology > DNA > DNA structure
Systems Biology > Epigenomics > Chromatin architecture
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.