- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Insertional Mutagenesis of Chlamydomonas reinhardtii
Published: Vol 5, Iss 24, Dec 20, 2015 DOI: 10.21769/BioProtoc.1680 Views: 9006
Reviewed by: Maria SinetovaIgor Cesarino Anonymous reviewer(s)
Original research article
The authors used this protocol in:

Nov 2014

Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Related protocols


Abstract
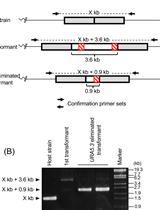
The unicellular microalga Chlamydomonas reinhardtii (C. reinhardtii) has been used as a reference model for numerous fields of research. Principle research areas are eukaryotic flagellar structure and function, basal bodies (centrioles), cell-cell recognition, cell cycle control, chloroplast biogenesis, phototaxis, nonphotochemical quenching, and especially photosynthesis for C. reinhardtii can grow in the dark on an organic carbon (e.g. acetate), and thus provides advantages over land plants (Harris, 2001; Peers et al., 2009). C. reinhardtii has a short life cycle, a sequenced genome (Merchant et al., 2007), and a growing molecular toolbox for forward and reverse genetic studies, including transformation protocols, gene silencing (Kim and Cerutti, 2009; Molnar et al., 2009), and fluorescent protein-tag (Rasala et al., 2013). There are two commonly used methods for C. reinhardtii transformation – electroporation and glass bead agitation. Electroporation is normally restricted to strains with cell wall, as it kills cell-wall-deficient strains effectively if without careful handling of osmosis. Electroporation also requires special instruments such as electroporator and cuvettes. In contrast, glass bead agitation uses simple lab equipment. The mild shear created by agitation in the presence of glass bead allows cell-wall-deficient strains to take up DNA. If glass bead method is to be applied to cell-wall strains, cells need to be treated with autolysin (http://www.chlamy.org/methods/autolysin.html) to partially lyse the wall components. A pitfall of both methods is that the DNAs are often shortened by nuclease once entering the cells, making the downstream PCR-based genotyping of insertion site rather difficult. Here I describe an improved design of insertional mutagenesis used in (Tsai et al., 2014), and the transformation protocol using glass bead as previously described in (Kindle, 1990) with minor modification. The putative mutants can be selected by autotrophic or antibiotic resistance markers, and the disrupted loci can be mapped by methods such as plasmid rescue (Peers et al., 2009) and SiteFinding PCR (Tan et al., 2005).
Materials and Reagents
- 15 and 50 ml conical tubes
- Petri dishes (90 mm in diameter)
- pHYG3 plasmid (http://chlamycollection.org/plasmid/phyg3/) or plasmid carrying other selection markers
- C. reinhardtii cell wall-less strain such as dw15 (cw15, nit1, mt+; http://chlamycollection.org/strain/cc-4619-cw15-nit1-mt-dw15-1/)
- Agar (Caisson Laboratories, Phytoblend)
- TAP medium (Harris, 1989)
- Restriction enzyme for linearizing the plasmid DNA, such as PvuII (New England Biolabs)
- Hygromycin (Life Technologies) or other selection means
- TAP agar plates (see Recipes)
- Top agar (see Recipes)
Equipment
- Common bench-top vortexer
- Spectrophotometer for optical density (OD) measurement, Z2 Coulter Counter (Beckman Coulter) or a hemocytometer
- 250 ml flask
- Bench-top centrifuge capable of centrifugation at 1,500 x g and accommodating 50 ml conical tubes
- Shaker
- Z2 Coulter Counter or hemocytometer
- Glass beads 425-600 μm (Sigma-Aldrich, catalog number: G8772 )
- Glass tubes or round bottom plastic tubes with or without cap (~10 mm in diameter)
Procedure
- DNA preparation (assumes use of pHYG3)
- Digest pHYG3 plasmid (http://www.biologie.uni-regensburg.de/Genetik/Mages/pHyg3.html) DNA with PvuII. PvuII was chosen specifically for pHYG3 plasmid because it releases a DNA fragment that contains the hygromycin B resistance gene aph7 with short flanking sequences. Based on our experience and others, linear DNAs are often shortened on each end after entering the cells of C. reinhardtii-ranging from dozens to hundreds of nucleotides-due to the exonuclease activity. It makes it harder to map the disrupted loci because the precise sequence of the insertion is unpredictable and different in each clone. Hence, we recommend using a DNA fragment that is just enough to cover the selectable marker (i.e. promoter, coding sequence, and terminator), so if the transformed DNAs are shortened to an extent that compromises the integrity of the marker, the clones harboring them will not be pulled out of the screen. In other words, the clones in the mutant library have the insertion whose length lies between the PvuII-digested fragment and the combined length of promoter, coding sequence, and terminator.
- Gel-excise the shorter, 2,012-bp fragments of the pHYG3 plasmid.
- Digest pHYG3 plasmid (http://www.biologie.uni-regensburg.de/Genetik/Mages/pHyg3.html) DNA with PvuII. PvuII was chosen specifically for pHYG3 plasmid because it releases a DNA fragment that contains the hygromycin B resistance gene aph7 with short flanking sequences. Based on our experience and others, linear DNAs are often shortened on each end after entering the cells of C. reinhardtii-ranging from dozens to hundreds of nucleotides-due to the exonuclease activity. It makes it harder to map the disrupted loci because the precise sequence of the insertion is unpredictable and different in each clone. Hence, we recommend using a DNA fragment that is just enough to cover the selectable marker (i.e. promoter, coding sequence, and terminator), so if the transformed DNAs are shortened to an extent that compromises the integrity of the marker, the clones harboring them will not be pulled out of the screen. In other words, the clones in the mutant library have the insertion whose length lies between the PvuII-digested fragment and the combined length of promoter, coding sequence, and terminator.
- Growth condition
In general, cells were grown in liquid TAP medium under continuous light (70-80 μmol/m-2 s-1) at 22 °C, with shaking at 100 rpm; or ambient room temperature (~22 °C) for solid media. - Glass bead transformation
- Grow cultures in TAP media (50 ml in 250 ml flask) until cell density reaches 1-2 x 106 cells/ml determined by Z2 Coulter Counter or hemocytometer, or approximately 0.2~0.3 OD550 by spectrophotometer.
- While cells are growing, prepare hardware: add ~0.3 g (~300 µl) glass beads to test tubes, autoclave.
- Pellet the cells by centrifugation (1,500 x g for 2 min), and resuspend in TAP to a concentration of ~2 x 107 cells/ml.
- Transfer 0.3 ml of cells to the glass bead-containing test tube, add ~1 µg linearized plasmid DNA prepared earlier (see Procedure A DNA preparation), and vortex 15 sec at top speed (Note 2).
- Add 6 ml TAP to the test tube, carefully transfer the cells (6 ml + 0.3 ml) to a new sterile 15 ml conical tube, and shake under growth conditions overnight (Note 3) at 100 rpm.
- After overnight recovery, add 6 ml of top agar (microwave to melt the top agar, cool to ~40 °C before pouring into the overnight culture) into the overnight culture (1:1 mixture). Mix gently.
- Immediately pour the 1:1 mixture onto TAP plates (6 ml for one regular Petri dish, 2 Petri dishes needed), let set (Note 4).
- Wrap plates with parafilm (cut slits to allow gas exchange), and place under lights. Colonies should start appearing in 5 to 7 days on the plate surface or embedded within the top agar layer. Generally speaking, 50 ml of culture (step C1) produces approximately 500 colonies after transformation. To create a mutant library ranging the entire C. reinhardtii genome (about 18,000 genes), it would require ~100 transformations in order to get a 3 fold coverage. Use sterile toothpicks or pipet tips to pick up the colonies.
- Grow cultures in TAP media (50 ml in 250 ml flask) until cell density reaches 1-2 x 106 cells/ml determined by Z2 Coulter Counter or hemocytometer, or approximately 0.2~0.3 OD550 by spectrophotometer.
Notes
- Transformation by glass beads is only applicable to cell wall-less strains (e.g. dw15, cw15) (Kindle, 1990). For the transformation of cell-wall strains, electroporation is more commonly used (https://www.thermofisher.com/order/catalog/product/A14258).
- Do not vortex the cells too vigorously otherwise DNA fragments tend to break, especially for longer fragments.
- If cell duplication during the overnight shaking (Procedure C, step C5) is a concern (e.g. to avoid duplicated clones for mutant screen), do 8-12 h shaking instead.
- Optional: Pre-warm the agar plates at 37 °C before pouring the 1:1 mixture (Procedure C, step C7).
Recipes
- TAP agar plates
0.8% agar with 10 μg/ml hygromycin - Top agar
TAP medium with 0.4% agar
Acknowledgments
This work was supported by the US Air Force Office of Scientific Research [Grant FA9550-11-1-0264 (to C. B.)], by a Strategic Partnership grant from the MSU Foundation (to C. B.), and by MSU AgBioResearch (C. B.).
References
- Harris, E. H. (2001). Chlamydomonas as a model organism. Annu Rev Plant Physiol Plant Mol Biol 52: 363-406.
- Kim, E. J. and Cerutti, H. (2009). Targeted gene silencing by RNA interference in Chlamydomonas. Methods Cell Biol 93: 99-110.
- Kindle, K. L. (1990). High-frequency nuclear transformation of Chlamydomonas reinhardtii. Proc Natl Acad Sci U S A 87(3): 1228-1232.
- Hoober, J. K. (1989). The Chlamydomonas sourcebook. A comprehensive guide to biology and laboratory use. Elizabeth H. Harris. Academic Press, San Diego, CA, 1989. xiv, 780 pp., illus. $145. Science 246(4936): 1503-1504.
- Merchant, S. S., Prochnik, S. E., Vallon, O., Harris, E. H., Karpowicz, S. J., Witman, G. B., Terry, A., Salamov, A., Fritz-Laylin, L. K., Marechal-Drouard, L., Marshall, W. F., Qu, L. H., Nelson, D. R., Sanderfoot, A. A., Spalding, M. H., Kapitonov, V. V., Ren, Q., Ferris, P., Lindquist, E., Shapiro, H., Lucas, S. M., Grimwood, J., Schmutz, J., Cardol, P., Cerutti, H., Chanfreau, G., Chen, C. L., Cognat, V., Croft, M. T., Dent, R., Dutcher, S., Fernandez, E., Fukuzawa, H., Gonzalez-Ballester, D., Gonzalez-Halphen, D., Hallmann, A., Hanikenne, M., Hippler, M., Inwood, W., Jabbari, K., Kalanon, M., Kuras, R., Lefebvre, P. A., Lemaire, S. D., Lobanov, A. V., Lohr, M., Manuell, A., Meier, I., Mets, L., Mittag, M., Mittelmeier, T., Moroney, J. V., Moseley, J., Napoli, C., Nedelcu, A. M., Niyogi, K., Novoselov, S. V., Paulsen, I. T., Pazour, G., Purton, S., Ral, J. P., Riano-Pachon, D. M., Riekhof, W., Rymarquis, L., Schroda, M., Stern, D., Umen, J., Willows, R., Wilson, N., Zimmer, S. L., Allmer, J., Balk, J., Bisova, K., Chen, C. J., Elias, M., Gendler, K., Hauser, C., Lamb, M. R., Ledford, H., Long, J. C., Minagawa, J., Page, M. D., Pan, J., Pootakham, W., Roje, S., Rose, A., Stahlberg, E., Terauchi, A. M., Yang, P., Ball, S., Bowler, C., Dieckmann, C. L., Gladyshev, V. N., Green, P., Jorgensen, R., Mayfield, S., Mueller-Roeber, B., Rajamani, S., Sayre, R. T., Brokstein, P., Dubchak, I., Goodstein, D., Hornick, L., Huang, Y. W., Jhaveri, J., Luo, Y., Martinez, D., Ngau, W. C., Otillar, B., Poliakov, A., Porter, A., Szajkowski, L., Werner, G., Zhou, K., Grigoriev, I. V., Rokhsar, D. S. and Grossman, A. R. (2007). The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science 318(5848): 245-250.
- Molnar, A., Bassett, A., Thuenemann, E., Schwach, F., Karkare, S., Ossowski, S., Weigel, D. and Baulcombe, D. (2009). Highly specific gene silencing by artificial microRNAs in the unicellular alga Chlamydomonas reinhardtii. Plant J 58(1): 165-174.
- Peers, G., Truong, T. B., Ostendorf, E., Busch, A., Elrad, D., Grossman, A. R., Hippler, M. and Niyogi, K. K. (2009). An ancient light-harvesting protein is critical for the regulation of algal photosynthesis. Nature 462(7272): 518-521.
- Rasala, B. A., Barrera, D. J., Ng, J., Plucinak, T. M., Rosenberg, J. N., Weeks, D. P., Oyler, G. A., Peterson, T. C., Haerizadeh, F. and Mayfield, S. P. (2013). Expanding the spectral palette of fluorescent proteins for the green microalga Chlamydomonas reinhardtii. Plant J 74(4): 545-556.
- Tan, G., Gao, Y., Shi, M., Zhang, X., He, S., Chen, Z. and An, C. (2005). SiteFinding-PCR: a simple and efficient PCR method for chromosome walking. Nucleic Acids Res 33(13): e122.
- Tsai, C. H., Warakanont, J., Takeuchi, T., Sears, B. B., Moellering, E. R. and Benning, C. (2014). The protein Compromised Hydrolysis of Triacylglycerols 7 (CHT7) acts as a repressor of cellular quiescence in Chlamydomonas. Proc Natl Acad Sci U S A 111(44): 15833-15838.
Article Information
Copyright
© 2015 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Tsai, C. and Benning, C. (2015). Insertional Mutagenesis of Chlamydomonas reinhardtii. Bio-protocol 5(24): e1680. DOI: 10.21769/BioProtoc.1680.
Category
Plant Science > Phycology > DNA > Mutagenesis
Molecular Biology > DNA > Mutagenesis
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.