

Colocalization Analysis for Cryosectioned and Immunostained Tissue Samples with or without Label Retention Expansion Microscopy (LR-ExM) by JACoP
JACoP对冷冻切片和免疫染色组织样本进行共定位分析,无论是否使用标签保留扩展显微镜 ( LR -ExM )
发布: 2022年03月05日第12卷第5期 DOI: 10.21769/BioProtoc.4336 浏览次数: 4790
评审: Nafisa M. JadavjiHai HuangKai Yuan

参见作者原研究论文
The authors used this protocol in:

Jun 2021
Advertisement









Abstract
Asymmetric cell division (ACD) is fundamental for balancing cell proliferation and differentiation in metazoans. During active neurogenesis in the developing zebrafish forebrain, radial glia progenitors (RGPs) mainly undergo ACD to produce one daughter with high activity of Delta/Notch signaling (proliferative cell fate) and another daughter with low Delta/Notch signaling (differentiative cell fate). The cell polarity protein partitioning-defective 3 (Par-3) is critical for regulating this process. To understand how polarized Par-3 on the cell cortex can lead to differential Notch activity in the nuclei of daughter cells, we combined an anti-Delta D (Dld) -atto 647N antibody uptake assay with label retention expansion microscopy (LR-ExM), to obtain high resolution immunofluorescent images of Par-3, dynein light intermediate chain 1 (Dlic1), and Dld endosomes in mitotic RGPs. We then developed a protocol for analyzing the colocalization of Par-3, Dlic1, and endosomal DeltaD, using JACoP (Just Another Co-localization Plugin) in ImageJ software (Bolte and Cordelières, 2006). Through such analyses, we have shown that cytosolic Par-3 is associated with Dlic1 on Dld endosomes. Our work demonstrates a direct involvement of Par-3 in dynein-mediated polarized transport of Notch signaling endosomes. This bio-protocol may be generalizable for analysis of protein co-localization in any cryosectioned and immunostained tissue samples.
Keywords: Par-3 (Par-3)Background
Asymmetric cell division (ACD) of progenitor/stem cells is critical for generating cellular diversity in tissue differentiation and organogenesis during development (Chenn and McConnell, 1995; Fishell and Kriegstein, 2003; Williams et al., 2011). During ACD, each daughter cell acquires distinct fates after mitosis by asymmetric inheritance of cellular components (so-called cell fate determinants), e.g., one continuing to self-renew and the other embarking on a differentiation path (Horvitz and Herskowitz, 1992; Knoblich, 2001). Previous studies from our laboratory and others have shown that cell polarity protein Par-3 is required to establish the asymmetric activity of Notch signaling between daughter cells (Notchhi vs. Notchlo) (Mizutani et al., 2007; Bultje et al., 2009; Coumailleau et al., 2009; Dong et al., 2012; Derivery et al., 2015; Kressmann et al., 2015), a key regulator of cell fate decisions (Artavanis-Tsakonas et al., 1999; Gaiano and Fishell, 2002). Recent studies show that Notch signaling in stem cell division is heavily controlled by endocytic trafficking (Chitnis, 2006; Daeden and Gonzalez-Gaitan, 2018). In Drosophila sensor organ precursor (SOP) cells, Notch signaling endosomes undergo directional movement (Derivery et al., 2015). Built on these previous findings, we investigated how Par-3 might regulate the dynamics of Dld endosomes in mitotic RGPs (Zhao et al., 2021).
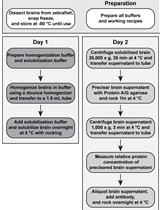
In this protocol, we describe our method for visualizing and analyzing cytoplasmic Par-3 in association with the dynein complex subunit Dlic1 and Dld endosomes in zebrafish RGP cells. The antibody uptake assay was used to label Dld endosomes in zebrafish embryos (Tong et al., 2019; Zhao et al., 2021). Following live labeling, cryosectioning, immunofluorescent staining, and label retention microscopy (LR-ExM) (Shi et al., 2021) were carried out. We have optimized these procedures and the protocol detailed here should be generalizable to all cryosectioned and immunostained tissue samples, not limited to zebrafish. Subsequently, we obtained high-resolution images of mitotic RGP cells in the processed samples, using spin-disk confocal microscopy. The cytoplasmic areas between two nuclei of the anaphase/telophase RGPs were designated as the regions of interest (ROIs) for colocalization analyses of Par-3, Dlic1, and Dld. Because the size of endosomes (average diameters of 250 nm to 1000 nm) is much larger than the pixel size of fluorescent images, the fluorescence signals colocalized on Dld endosomes may not appear to exactly overlap with one another. Based on previously published reports (Bolte and Cordelières, 2006; Derivery et al., 2015), we developed the method for measuring co-localization in the context of endosomes in this protocol. For all three channels, we measured the colocalization between every two fluorescent channels in the form of Manders’ Coefficients (i.e., the proportion of each fluorescence colocalized with another) by using the JACoP plugin in ImageJ (Manders et al., 1993). Our results uncovered that Par-3 is colocalized with Dlic1 on Dld endosomes, hence revealing a direct role of cytoplasmic Par-3 in actively localizing intracellular determinants during ACD of RGP cells (Zhao et al., 2021).
Materials and Reagents
FisherbrandTM SuperfrostTM Plus microscope slides (Fisher Scientific, catalog number: 22-037-246)
6-well glass bottom plates (CellVis, catalog number: P06-1.5H-N)
Zebrafish strains wild-type zebrafish embryos were obtained from natural spawning of AB adults, staged and maintained according to established protocols (Kimmel et al., 1995)
Zebrafish embryos were raised at 28.5 °C in 0.3× Danieau’s embryo medium (Zhao et al., 2021)
Capillaries, 1.2 mm OD, 0.9 mm ID, with filament (World Precision Instruments, catalog number: 1B120F-6)
Tricaine (Sigma-Aldrich, CAS Number: 886-86-2, catalog number: E10521)
Phenol red (Sigma-Aldrich, catalog number: P0290)
Primary antibodies used in this study:
Mouse anti-Dld (Abcam, catalog number: ab73331; RRID, catalog number: AB_1268496; lot, catalog number: GR115501-3, 1:200 dilution for immunostaining)
Chicken anti-GFP (Abcam, catalog number: ab13970; RRID:AB_300798, lot GR3190550-20, 1:500 dilution for immunostaining)
Rabbit anti-Par-3 (Millipore 07-330; RRID:AB_2101325; lot 3322358, 1:500 for immunostaining)(validated in this study)
Guinea pig anti-DLIC1-Cter (a gift from Dr. T. Uemura, 1:100 for immunostaining) (Satoh et al., 2008)
Secondary antibodies used for immunofluorescent labeling:
Alexa®-conjugated goat anti-rabbit (Alexa 568, Invitrogen, catalog number: A11011, RRID. AB_143157, lot 792518, 1:2,000 dilution)
Goat anti-chicken (Alexa 488, Invitrogen, catalog number: A11039, RRID:AB_142924, lot 2020124, 1:2,000 dilution)
Goat anti-mouse (Alexa 488, Invitrogen, catalog number: A11002, RRID:AB_2534070, lot 1786359, 1:2,000 dilution)
Goat anti-guinea pig (Alexa 488, Invitrogen, catalog number: A11073, RRID:AB_2534117, lot 46214A, 1:2,000 dilution) or Donkey anti-guinea pig (Alexa 647, Jackson Labs, catalog number: 706-605-148, RRID:AB_2340476, lot 102649-478, 1:2,000 dilution)
Anti-Mouse-IgG-Atto647N (Sigma-Aldrich, catalog number: 50185, 1 mg/mL)
Secondary antibodies used for tri-functional linker conjugation in LR-ExM were:
Goat anti-Guinea Pig IgG (H+L) unconjugated secondary antibody (Invitrogen, catalog number: A18771, RRID:AB_2535548)
Goat anti-Rabbit IgG (H+L) Cross-Adsorbed unconjugated secondary antibody (Invitrogen, catalog number: 31212, RRID:AB_228335)
Goat anti-Chicken IgY (H+L) unconjugated secondary antibody (Invitrogen, catalog number: A16056, RRID:AB_2534729)
NHS-MA-Biotin conjugated anti-Chicken IgY, NHS-MA-Biotin conjugated anti-Rabbit IgG, and NHS-MA-DIG conjugated anti-Guinea Pig IgG (Gift from Dr. Shi) (Shi et al., 2021)
DAPI solution (1 mg/mL) (Thermo ScientificTM, catalog number: 62248)
Tween 20 (Sigma-Aldrich, CAS Number: 9005-64-5, catalog number: P-9416)
TritonTM X-100 (Sigma-Aldrich, CAS Number: 9036-19-5)
Dimethyl sulfoxide (DMSO) (Sigma-Aldrich, CAS Number: 67-68-5)
Natural goat serum (Invitrogen, catalog number: 31872)
Glycerol (Sigma-Aldrich, CAS Number: 56-81-5G, catalog number: 9012)
Phosphate-buffered saline (PBS) 10× solution, pH 7.4 (GibcoTM, catalog number: 70011044)
Sucrose (Sigma-Aldrich, CAS number: 57-50-1)
Tissue-PlusTM O.C.T. compound (Fisher Scientific, catalog number: 23-730-571)
UltraPureTM low melting point agarose (Invitrogen, catalog number: 16520050)
Sodium acrylate (Sigma-Aldrich, CAS Number: 7446-81-3)
Acrylamide (Sigma-Aldrich, CAS Number: 79-06-1, catalog number: A9099)
Sodium chloride (Sigma-Aldrich, CAS Number: 7647-14-5, catalog number: 204439)
N,N,N′,N′-Tetramethyl ethylenediamine, TEMED (Sigma-Aldrich, CAS Number: 110-18-9, catalog number: 1.10732)
Ammonium persulfate, APS (Thermo ScientificTM, catalog number: 17874)
Proteinase K (Sigma-Aldrich, CAS Number: 39450-01-6, catalog number: P4850)
N,N′-Methylenebis-acrylamide (Sigma-Aldrich, CAS Number: 110-26-9, catalog number: M7279)
30× Danieau’s embryo medium (see Recipes)
Blocking buffer for immunostaining (see Recipes)
Washing buffer (see Recipes)
LR-ExM gelation buffer and digestion buffer (see Recipes)
Gelation buffer (see Recipes)
Monomer solution (see Recipes)
Digestion buffer (see Recipes)
Post-digestion staining buffer (see Recipes)
Equipment
Stero dissection microscope (Zeiss Stemi 2000 with a maximum magnification of 50)
Air pressure injector( Narishige IM 300 microinjector)
Micromanipulators (WPI M3301R, World Precision Instruments, Sarasota, FL, USA)
Flaming-Brown P897 needle puller (Sutter Instruments, Novato, CA, USA)
Nikon CSU-W1 Spinning Disk/confocal microscope
Leica CM1850 Cryostat
Software
Micro-Manager 2.0 gamma (μManagerTM, University of California), https://micro-manager.org/
Fiji (Schindelin et al., 2012), https://imagej.net/software/fiji/
Prism 9, GraphPad, https://www.graphpad.com/
Procedure
文章信息
版权信息
© 2022 The Authors; exclusive licensee Bio-protocol LLC.
如何引用
Readers should cite both the Bio-protocol article and the original research article where this protocol was used:
- Zhao, X., Garcia, J., Royer, L. A. and Guo, S. (2022). Colocalization Analysis for Cryosectioned and Immunostained Tissue Samples with or without Label Retention Expansion Microscopy (LR-ExM) by JACoP. Bio-protocol 12(5): e4336. DOI: 10.21769/BioProtoc.4336.
- Zhao, X., Garcia, J. Q., Tong, K., Chen, X., Yang, B., Li, Q., Dai, Z., Shi, X., Seiple, I. B. and Huang, B. (2021). Polarized endosome dynamics engage cytoplasmic Par-3 that recruits dynein during asymmetric cell division. Sci Adv 7(24): eabg1244.
分类
细胞生物学 > 细胞成像 > 荧光
神经科学 > 发育 > 免疫荧光
分子生物学 > 蛋白质 > 蛋白质-蛋白质相互作用
您对这篇实验方法有问题吗?
在此处发布您的问题,我们将邀请本文作者来回答。同时,我们会将您的问题发布到Bio-protocol Exchange,以便寻求社区成员的帮助。