
- Home
- Protocols

-

Pairwise interactions between microbial species
Last updated date: May 26, 2023 Views: 926 Forks: 0
To measure the strength of pairwise interactions in the experiments, we randomly chose six isolates—selected from the different genera Leuconostoc, Pseudomonas, Yersinia, Pantoea, Klebsiella, and Acinetobacter—from the bacterial library. We first measured the carrying capacity
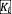
(i.e., the species abundance at equilibrium in the absence of any competitor species) of these isolates through exposing them to 7 cycles of daily dilutions in monoculture, followed by plating and colony counting at the end of the 7th cycle. We then co-cultured 15 pairs (all possible combinations) of these isolates over 7 dilution cycles and measured the species abundance at the end point (via sample dilution and colony counting on agar plates). Together with the measured carrying capacities, these species abundances were used to assess the strength of interactions via the relationship

, which can be easily derived from the gLV model. Table S1 shows that all pairs of isolates coexist under low nutrient concentrations (
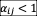
for all the experimentally measured interactions). For higher nutrient concentrations, we considered 2 initial relative abundance for each pair of species (initial species ratios 95:5 and 5:95, measured via culture volume), which allows to identify cases of bistability in which either species can lead its competitor to extinction. For coexisting pairs, the value of
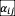
was calculated as stated above. In cases of competitive exclusion (species i always drives species j to extinction), we inferred that
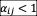
and

. For bistability (the high-abundance species drives the low-abundance one to extinction), we inferred that
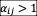
and

. Tables S2 and S3 show the measured interaction matrices under medium and high nutrients concentrations. We found the interaction matrices measured in the experiment are densely connected matrices (Tables S1-S3), which means that
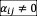
for most (or all) the species interactions. This result is consistent with previously observed microbial community interaction networks (7) and supports the assumption a dense interaction matrix in our theoretical model.
Ecological communities, including microbial communities different from the ones in our experiments, need not to be densely connected. Our model can account for this fact through incorporating the average connectance C (the fraction of non-zero interactions in the interaction network) in the stability criteria (16). In this way, the main effect of network connectance in community dynamics is equivalent to replacing S by SC on the horizontal axis of the phase diagrams in Figs. 1E and 1F (17). Therefore, a finite fraction of zero (or negligibly small) interaction strengths does not qualitatively change the phases of community dynamics and their relative positions on the phase diagram.
Related files
 Reading2.pdf
Reading2.pdf - Hu, J and Gore, J(2023). Pairwise interactions between microbial species. Bio-protocol Preprint. bio-protocol.org/prep2322.
- Hu, J., Amor, D. R., Barbier, M., Bunin, G. and Gore, J.(2022). Emergent phases of ecological diversity and dynamics mapped in microcosms. Science 378(6615). DOI: 10.1126/science.abm7841
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.