
- Home
- Protocols

-

Identifying antigenic switching by clonal cell barcoding and nanopore sequencing in trypanosomes
Last updated date: Aug 4, 2023 Views: 812 Forks: 0
Identifying antigenic switching by clonal cell barcoding and nanopore sequencing in trypanosomes
Abdoulie O. Touray1,2, Tamara Sternlieb1, Tony Isebe2 and Igor Cestari1,2,*
1Institute of Parasitology, McGill University, Ste Anne de Bellevue, QC H9X 3V9, Canada
2Division of Experimental Medicine, McGill University, Montreal, QC, H4A 3J1, Canada
*For correspondence: igor.cestari@mcgill.ca
Abstract
Many organisms alternate the expression of genes from large gene sets or gene families to adapt to environmental cues or immune pressure. The single-celled protozoan pathogen Trypanosoma brucei spp. periodically change their homogeneous surface coat of variant surface glycoproteins (VSGs) to evade host antibodies during infection. They express one out of ~2,500 VSG genes at a time from telomeric expression sites (ESs) and periodically change their expression by transcriptional switching or recombination. Attempts to track VSG switching have previously relied on genetic modifications of ES sequences with drug-selectable markers or genes encoding fluorescent proteins. However, genetic modifications of the ESs can interfere with the binding of proteins that control VSG transcription and/or recombination, thus affecting VSG expression and switching. Other approaches include Illumina sequencing of the VSG repertoire, which shows VSGs expressed in the population rather than cell switching, and the short reads often limit distinguishing the large set of VSG genes. Here we describe a methodology to study antigenic switching without modifications of the ES sequences. Our protocol enables the detection of VSG switching at nucleotide resolution using multiplexed clonal cell barcoding to track cells and nanopore sequencing to identify cell-specific VSG expression. We also developed a computational pipeline that takes DNA sequences and outputs VSG expressed by cell clones. This protocol can be easily adapted to study clonal cell expression of large gene families in prokaryotes or eukaryotes.
Keywords: Oxford nanopore sequencing, trypanosomes, switching, DNA barcode, VSG-seq, variant surface glycoproteins, antigenic variation
This protocol is used in: eLife (2023), DOI: 10.7554/eLife.89331.1
Graphical overview
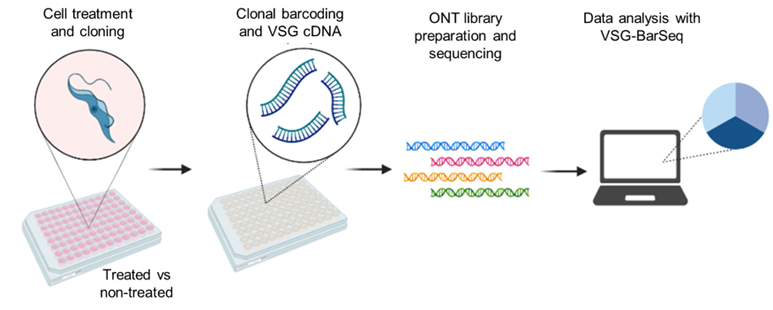
Background
Trypanosoma brucei is a single-celled protozoan parasite that causes African trypanosomiasis and evades the host antibody response changing its surface coat by antigenic variation (Cestari and Stuart, 2018). T. brucei expresses a single variant surface glycoprotein (VSG) gene from one of the 20 telomeric expression sites (ESs) and periodically switches to a different VSG via transcriptional switching between ESs or by VSG gene recombination. T. brucei has an extensive repertoire of over 2,500 VSG genes and pseudogenes located in telomeric and sub-telomeric regions of large chromosomes. VSG genes are also found on dozens of mini chromosomes often used for VSG gene recombination. VSG genes are about 2 kb in length with conserved C-terminus sequences. VSG recombination can occur by gene or segmental gene conversion, resulting in new mosaic VSG sequences. The mechanisms controlling VSG monogenic expression and switching likely entail multiple processes, including the control of VSG repression and expression via proteins associated with telomeric ESs. Several proteins associate with the telomeric repeats or ESs to regulate VSG gene expression and/or switching, such as the repressor activator protein 1 (Touray et al., 2023), phosphatidylinositol 5-phosphatase (Cestari et al., 2019), telomeric repeat-binding factor (Jehi et al., 2014), VSG exclusion protein 2 (Faria et al., 2019), and ES body 1 protein (Lopez-Escobar et al., 2022); for a review on additional proteins controlling VSG expression and switching see (Cestari and Stuart, 2018).
Approaches used to study VSG switching rely on genetic modifications that disrupt the ES DNA sequences by incorporating drug-selectable markers or fluorescent proteins downstream of the promoter sequence and upstream of the VSG gene (Aitcheson et al., 2005; Rudenko et al., 1998; Ulbert et al., 2002), or by adding exogenous endonuclease sites resulting in DNA breaks (Boothroyd et al., 2009; Glover et al., 2013). However, the ES modifications might disrupt protein binding sites and thus affect VSG switching rates, e.g., RAP1 binds to 70 bp and telomeric repeats flanking ES VSGs genes and repress their transcription, and disruption of its binding dramatically increases VSG switching rates (Touray et al., 2023). In addition, the genetic modifications of ESs are laborious and restrict the use of drug-selectable markers available for other genetic changes, such as gene knockout or expression of mutant variants in the cell. Other studies used Illumina RNA-seq to investigate VSG expression at a population level (Mugnier et al., 2015). Although this approach helps identify expressed VSG genes, many short-reads fail to align uniquely to the genome and unambiguously identify and distinguish VSG genes expressed from the extensive repertoire of VSG genes/pseudogenes. Moreover, it does not identify switching cells but expressed VSG genes in the population.
Hence, we sought to develop an approach to track cell-specific antigenic switching without genetically modifying ES sequences in an adaptable high or medium-throughput fashion. We devised a method to detect VSG switching at nucleotide resolution using clonal cell barcoding and nanopore sequencing. We combine DNA barcoding to identify parasite cell clones and thus track switching and non-switching cells, and a broad-spectrum VSG primer for cDNA synthesis to capture all transcribed VSGs in parasite clones. Barcoded samples are multiplexed, and VSG cDNAs are sequenced using Oxford nanopore sequencing, followed by sequencing data analysis using a streamlined VSG-BarSeq pipeline to identify switchers. We performed VSG-BarSeq after a temporary knockdown of PIP5Pase, an enzyme that regulates VSG switching in T. brucei (Touray et al., 2023). We found that 99% of the clones switched VSG genes, whereas no switching was detected in the control non-knockdown cell line. The long nanopore reads helped identify complete VSG sequences and thus track modes of switching. We detected VSG switching by transcriptional and recombination mechanisms, indicating the approach's usefulness in studying antigenic variation. This protocol will enable the efficient study of VSG switching without genetic alterations of the ES. The protocol may be easily adapted to other organisms to study antigenic switching (e.g., Plasmodium, Giardia) or clonal expression analysis of large gene families in prokaryotes or eukaryotic cells.
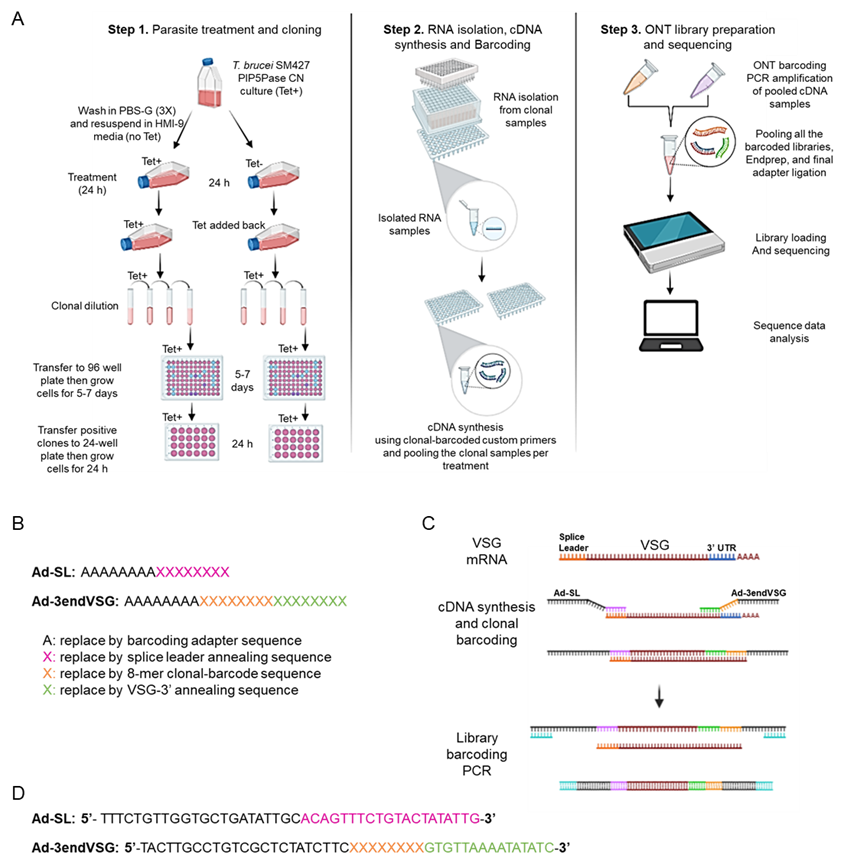
Figure 1. A schematic representation of clonal cell barcode and nanopore sequencing protocol. (A) Diagram of the clonal cell barcoding and nanopore sequencing workflow showing (step 1) the parasite treatment and cloning, (step 2) RNA isolation from the individual clones, cDNA synthesis, and clonal barcoding, and (step 3) ONT library preparation, sequencing, and data analysis using VSG-BarSeq script. (B) Scheme of the forward Adaptor Splice Leader (Ad-SL) primer and the reverse Adaptor-barcode-3’-end VSG primer (Ad-3endVSG) used for cDNA synthesis and clonal cell barcoding. (C) Diagram describing the cDNA synthesis, clonal barcoding, ONT library barcoding, and PCR amplification. (D) Sequences of the Ad-SL and Ad-3endVSG annealing primers used for cDNA synthesis and clonal cell barcoding (see the annealing regions in C). The sequences in black correspond to ~20 bp nanopore barcode adapter sequences, and the sequences in pink and green correspond to the forward Ad-SL and reverse Ad3endVSG primers, respectively. The Ad-SL pairs to the mRNA splice leader sequence while the reverse Ad3endVSG primer pairs to the conserved 3’-end of VSG mRNAs. The 8-nucleotide long variable sequence (clonal barcode) unique to each clone is shown in orange and depicted here as X. A complete list of primers is available in Supplementary Information.
Materials and Reagents
Biological materials
Trypanosoma brucei bloodstream form parasites.
Reagents
M-MuLV Reverse Transcriptase (New England Biolabs Ltd, catalog number: M02535)
M-MuLV Reverse Transcriptase 10X Buffer (New England Biolabs Ltd, catalog number: B02535)
MgCl2 (Invitrogen, catalog number: R0971)
Mag-Bind® Total Pure NGS (Omega Bio-Tek, catalog number: M1378-01)
NEBNext® End Repair Module (New England Biolabs Ltd, catalog number: E6050S)
Monarch® DNA Elution Buffer (New England Biolabs Ltd, catalog number: T1016L)
Native Barcoding Expansion 1-12 (Oxford Nanopore Technologies, catalog number: EXP-PBC001)
Blunt/TA Ligase Master Mix (New England Biolabs Ltd, catalog number: M0367S)
Taq DNA Polymerase with ThermoPol Buffer (New England Biolabs Ltd, catalog number: M0267S)
Deoxyribonucleotide triphosphate (dNTPs) mixture 10mM (Biobasic, catalog number: DD0056)
Agarose (Bishop Canada Inc., catalog number: AGA002.250)
Iscove's Modified Dulbecco's Medium (IMDM) powder (Life Technologies, catalog number: 12200069)
Sodium bicarbonate (Fisher Scientific, catalog number: S233-3).
Hypoxanthine (Millipore, catalog number: 4010CBC-25GM).
Sodium pyruvate (Wisent Inc., catalog number: 600-110-EL).
Bathocuproin sulfonate disodium salt hydrate (Thermo Scientific, catalog number: B22550.MD).
L-cysteine hydrochloride monohydrate (Thermo Scientific, catalog number: J14035-22).
2-mercaptoethanol (Sigma, catalog number:M722).
Penicillin Streptomycin (MP Biomedicals, catalog number:1670049).
Heat inactivated fetal bovine serum (FBS) (Life Technologies, catalog number:12484-028).
Neomycin (G418) stock solution (20 mg/ml) in 50 ml (Sigma, catalog number: N6386).
Tetracycline Hydrochloride (Fisher Scientific, catalog number: BP912-100)
Sodium phosphate dibasic anhydrous, Na2HPO4 (Fisher Scientific, catalog number: S374-500).
Sodium phosphate monobasic anhydrous, NaH2PO4 (Sigma, catalog number: S-0751).
Sodium chloride, NaCl (Fisher Scientific, catalog number: S-271-3).
Glucose (Sigma, catalog number: G5400-250G).
Sodium citrate (dihydrate) (Fisher Scientific, catalog number: S279-500).
Bovine serum albumin (BSA) (Biobasic, catalog number: A500023-0100).
Glycerol (Fisher Bioreagents, catalog number: BP229-1).
Tris base (Fisher Scientific, catalog number: BP152-5).
Glacial acetic acid (Fisher Chemical, catalog number: A38-212).
Ethylenediaminetetraacetic acid (EDTA) (Fisher Scientific, catalog number: BP120-500).
Trypan Blue Solution, 0.4% (Amresco, catalogue number K940-100ML).
Solutions
HMI-9 cell culture medium (see recipes)
Drug selectable markers (see recipes)
PBS-Glucose (see recipes)
Bloodstream stabilate freezing solution (see recipes)
50X Tris Acetate Ethylenediaminetetraacetic acid (EDTA) solution (see recipes)
Recipes
1. HMI-9 cell culture medium (1000 mL)
Reagent | Final concentration | Quantity or Volume |
Iscove's Modified Dulbecco's Medium (IMDM) | n/a | 17.7 g |
Sodium bicarbonate | 0.035 M | 3 g |
Hypoxanthine | 10 mM | 10 mL |
Sodium pyruvate | 100 mM | 10 mL |
Bathocuproine sulfonate disodium salt hydrate | 49.95 µM | 28.2 mg |
L-cysteine hydrochloride monohydrate | 1.04 mM | 182 mg |
2-mercaptoethanol | 14.3 M | 14 µL |
Nanopure MilliQ water | n/a | 879.9 mL |
Total volume | n/a | 900 mL |
Dissolve the IMDM and salts in 500 mL of water, and stir to mix thoroughly. Add 10 mL of hypoxanthine, 10 mL of sodium pyruvate, and 14 µL of 2-mercaptoethanol. Adjust water volume to 900 mL. Filter the medium using a 0.22 µm filter system of 250 mL. Add 100 units of sterile penicillin and 100 µg/mL of sterile streptomycin (Optional). Complete the media by adding 10% heat-inactivated fetal bovine serum (FBS). Store it at 4°C.
2. Drug-selectable markers
a. Neomycin (G-418) Stock solution (20 mg/mL)
Reagent | Final concentration | Quantity or Volume |
Neomycin (G-418) | n/a | 1 g |
Nanopure MilliQ water | n/a | 50 mL |
Total | n/a | 50 mL |
b. Tetracycline Stock solution (5 mg/mL)
Reagent | Final concentration | Quantity or Volume |
Tetracycline Hydrochloride | n/a | 0.5 g |
Nanopure MilliQ water | n/a | 10 mL |
Total | n/a | 10 mL |
Dissolve the drugs in water. Filter the dissolved drug solutions using a 0.22 µm syringe filter. Prepare 1 mL aliquots into 1.5 mL Eppendorf tubes in the hood, and store the aliquots at -20°C freezer.
3. Phosphate-buffered saline- Glucose (PBS-G) (1000 mL)
Reagent | Final concentration | Quantity or Volume |
Sodium phosphate dibasic anhydrous | 0.01 M | 1.42 g |
Sodium phosphate monobasic anhydrous | 0.01 M | 1.20 g |
Sodium chloride | 0.145 M | 8.5 g |
D-Glucose | 0.006 M | 1.081 g |
Nanopure MilliQ water | n/a | 1000 mL |
Total | n/a | 1000 mL |
Dissolve salts in 900 mL of water. Add D-Glucose and stir thoroughly to dissolve it. Adjust the pH to 7.0, and filter sterilize using a 0.22 µm filter.
4. Bloodstream stabilate freezing solution (500 mL)
Reagent | Final concentration | Quantity or Volume |
D-Glucose | 0.052 M | 9.3 g |
Sodium chloride | 0.036 M | 2.1 g |
Sodium citrate | 0.003 M | 0.75 g |
Bovine serum albumin (BSA) | n/a | 0.5 g |
Glycerol | n/a | 75.0 g |
Nanopure MilliQ water | n/a | 500 mL |
Total | n/a | 500 mL |
Dissolve the salts in 300 mL of water. Add D-glucose and stir thoroughly to dissolve it. Weight the volume of glycerol equivalent to 75 g in a separate container and add it to the solution. Adjust the volume of the solution to 500 mL, and stir thoroughly. Filter in 0.22 µm filter. Store it at 4°C.
5. 50X Tris Acetate Ethylenediaminetetraacetic acid (EDTA) solution (500 mL)
Reagent | Final concentration | Quantity or Volume |
Tris base | 1 M | 121 g |
Glacial acetic acid | 17.4 M | 28.6 mL |
Ethylenediaminetetraacetic acid (EDTA) | 0.5 M | 50 mL |
Nanopure MilliQ water | n/a | 421.4 mL |
Total volume | n/a | 500 mL |
Prepare a stock solution of 0.5 M EDTA in a separate tube by dissolving 18.6 g in 80 mL of water. Adjust the pH to 8.0 by adding 10 M Sodium hydroxide (NaOH) solution. Adjust the volume to 100 mL. Dissolve the Tris base in 300 mL water, add 28.6 mL of 17.4 M Glacial acetic acid and 50 mL of 0.5 M EDTA. Adjust the volume of the solution to 500 mL with water. Autoclave the solution. Store it at room temperature.
Laboratory supplies
Tissue culture flasks
25 cm2 TC-treated T-flask with filter cap (Biobasic, catalog number: SP81136)
75 cm2 TC-treated T-flask with filter cap (Biobasic, catalog number: SP81186)
Cell culture plates
96-well cell culture plates (WUXI NEST Biotechnology Co., catalog number: 101722BL01)
24-well plates TC plate, treated (Biobasic, catalog number: SP41135)
Sterile pipette tips
1000 µL pre-sterile barrier tips (Neptune, catalog number: BT100.96)
200 µL pre-sterile barrier tips (Neptune, catalog number: BT200)
10 µL pre-sterile barrier tips (Neptune, catalog number: BT10)
96-well plate bacterial total RNA mini-prep super kit (Biobasic, catalog number BS585-5)
96-well 2 mL Deep Plate, Natural, Edge Filled, 10 Plates/Bag (Biobasic, catalog number: BR581-96NS)
96-well Deep Collection Plates (Biobasic, catalog number: 107-E627LA2221)
Plate seals (Biobasic, catalog number: BS585-5)
96-well PCR plate (Life Technologies, catalog number: 4346907)
Foil sealing film, non-sterile (Celltreat Scientific Products, catalog number: 501535152)
Multichannel pipette (Eppendorf, catalog number: 4056991)
Heating block (Eppendorf, catalog number: 535028642)
Filter system 250 mL 0.2 µm pes (Fisher Scientific, catalog number: FB12566502).
Equipment
Ultrafocused-sonicator (Covaris, M220, catalog number: 006168)
Magnetic rack for 1.5 mL tubes (Promega, catalog number: PR-Z5342)
T100 Thermal Cycler (Bio-Rad, catalog number: 1861096)
Avanti J-E Centrifuge (Beckman Coulter, catalog number: JSE02M13)
AllegraTM 25R Centrifuge, TJ-25 Rotor (Beckman Coulter, catalog number: AJC024001)
Nanodrop (Nanodrop Spectrophotometer, catalog number: ND-1000)
Microcentrifuge (Eppendorf, catalog number: EP5401000137)
Water System Ultrapure (Nanopure MilliQ water) (Millipore Synergy, catalog number: F1CA45528 A)
Electrophoresis unit: Thermo Scientific Power Supply 400mA 300V (Fisher, catalog number: S65533Q)
Biological safety cabinet (NuAire Biological Safety Cabinet- Class II Type AIB3, catalog number: NU-425-300)
CellDrop™ Automated Cell Counter (DeNovix Cell Drop FL Fluorescence Cell Counter)
HEPA CO2 Incubator (Thermo Electron Corporation; Forma Series II, Water Jacketed, catalog number: 308606-30529)
Microscope (Nikon Eclipse TS100, catalog number: 302115)
-80 °C Freezer (New Brunswick Ultra-Low Temperature Freezer U101 Inova, catalog number: U9420-0000)
Software and datasets
Minimap2 (Li, 2018) (https://github.com/lh3/minimap2)
Samtools (Danecek et al., 2021) (https://www.htslib.org/)
DeepTools (Ramirez et al., 2016) (https://deeptools.readthedocs.io/en/develop/index.html)
Subread (Liao et al., 2014) (http://subread.sourceforge.net/featureCounts.html)
Rcgrep (https://github.com/dib-lab/rcgrep)
Integrative Genomics Viewer (IGV) (https://software.broadinstitute.org/software/igv/)
VSG-BarSeq (this work) (https://github.com/cestari-lab/VSG-Bar-seq)
Procedure
A. Parasite treatment and cloning
We recommend determining cell treatment conditions before starting this protocol. The conditions used in this protocol were optimized for T. brucei bloodstream forms of the 427 strain or conditional null (CN) cells derived from the single-marker 427 strain (Cestari and Stuart, 2015). The treatment described here is the knockdown for 24h of the T. brucei gene encoding PIP5Pase, which results in high rates of VSG switching (Touray et al., 2023). The PIP5Pase CN cell line is grown in G418 and phleomycin to maintain the selection of the tetracycline-inducible system. Tetracycline is added to induce expression of the PIP5Pase gene under the control of a procyclin promoter and a tetracycline operator.
Grow 5 mL of T. brucei cells seeded at 1.0x104 cells/mL in HMI-9 medium supplemented with 2 µg/mL of G418, 2.5 µg/mL of phleomycin, and 500 ng/mL of tetracycline in a 37°C incubator with 5% CO2 for 24h or until it reaches mid-log growth (~1.0x106 cells/mL). Throughout this protocol, cell growth will be as described above unless otherwise stated. Cells doubling time should be about 5.5-6h and viability about 90-95%. Avoid overgrowing cell culture (>1.5x106 cells/mL) because it will affect cell viability.
Transfer the 5 mL cell culture to a 15 mL Falcon tube and centrifuge at 3500 x g for 5 min at room temperature (RT). Discard the supernatant.
Resuspend the pellet in 10 mL PBS-glucose (PBS-G) pre-warmed at 37°C, then centrifuge the cells as in step A2. Discard the supernatant.
Repeat step A3 three times to ensure complete removal of tetracycline.
Split the cells in two 5 mL cell culture flasks (treated and non-treated groups) seeding each culture at 1.0x104 cells/mL in HMI-9 medium with 2 µg/mL G418 and 2.5 µg/mL of phleomycin.
Add 500 ng/mL of tetracycline to the non-treated flask (Tet +, control) and no tetracycline to the treatment flask (Tet -, knockdown) and grow the cells for 24h.
Add 500 ng/mL of tetracycline to the treatment flask (Tet -, knockdown). Quantify the cell concentration and viability by mixing 10 µL of cell culture and 10 µL of 0.4% Trypan blue staining. Add 10 µL to the CellDrop cell counter to obtain viability and cell concentration. Cell concentration should be about 1.0x105 cells/mL, and viability should be > 90%.
Add 9 mL of HMI-9 medium supplemented with 500 ng/mL of tetracycline to a 50 mL Falcon tube. Repeat the procedure to have three flasks for the treatment group and three for the non-treatment group.
From a starting cell concentration of 1.0x105 cells/mL (from A7 above), gently mix the cells by flicking the flasks and transfer 1 mL of the culture to the Falcon tube-1 containing 9 mL of HMI-9/FBS media (1 in 10 dilutions) to obtain a cell concentration of 1x104 cells/mL.
From the Falcon tube 1 (1x104 cells/mL) culture, transfer 1 mL to Falcon tube 2 to obtain a cell concentration of 1x103 cells/mL (1 in 10 dilutions). Repeat the same for Falcon tube 3 to obtain a cell concentration of 1x102 cells/mL.
Transfer 7 mL of the culture from Falcon tube 3 to a 75 cm2 cell culture flask containing 63 mL of HMI-9 medium supplemented with 2 µg/mL G418, 2.5 µg/mL of phleomycin, and 500 ng/mL of tetracycline to obtain a cell concentration of 10 cells/mL (1 in 10 dilutions).
Transfer 60 mL of the diluted cell culture from A11 to a 75 cm2 cell culture flask and add 140 mL of HMI-9 medium supplemented with 2 µg/mL G418, 2.5 µg/mL of phleomycin, and 500 ng/mL of tetracycline (3 in 10 dilutions) to obtain a final cell concentration of 3 cells/mL.
Aliquot the diluted parasite culture (3 cells/mL) onto ten 96-well plates (10 plates per treatment group), transferring 200 µL to each well such that the probability of obtaining one single cell per well (200 µL) is about 30%, i.e., one cell per well for a third of wells of a 96-well plate.
Grow the cells in 96-well plates in an incubator for 5 to 7 days.
Check the 96-well plate under a microscope to identify parasite clones. About 30% of the wells should contain parasites. (See Note 1)
Transfer 200 µL of parasite clones onto 24-well plates and add 1.8 mL of fresh HMI-9 with 2 µg/mL G418, 2.5 µg/mL of phleomycin, and 500 ng/mL of tetracycline.
Grow the clones in the 24-well plates for 24 h to increase the number of cells for RNA extraction.
(Optional) Aliquot 600 µL of each clonal population onto new 96 deep-well collection plates and add 600 µL of Freezing Solution. Freeze the parasites at -80o C for short-term storage (2-3 weeks) or liquid nitrogen for long-term storage.
The remaining 1.4 mL of the clonal parasite cultures are used for RNA extraction.
B. RNA extraction
Transfer 1.4 mL of each clonal culture (about 1.4 x106 cells) into 96 deep-well collection plates, centrifuge at 3500 x g for 5 min at room temperature (RT), and pour off the supernatant. (See Note 2)
Add 350 µL buffer Rlysis-BG to each well and resuspend the cells by pipetting up and down.
Thoroughly seal the plate with a sealing film to prevent cross-contamination of the samples, and immediately mix the samples by inverting the plate three times.
Briefly spin the plate at 1000 x g for 30 seconds to collect the solution to the bottom of the wells.
Add 175 µL of ethanol to each well, tightly seal the plate with a new sealing film, and mix thoroughly by inverting three times.
Place the EZ-10 96-Well Plate (filtration column plate provided in the 96-well plate bacterial total RNA mini-preps super kit) on top of a new Deep Well Collection Plate and transfer the lysate from step 5 into the columns on the EZ-10 96-Well Plate.
Centrifuge the plate at 5,000 x g for 2 min at RT and discard the flow through.
Placethe EZ-10 96-well plate back on the Deep Well Collection Plate and add 500 µL of universal GT solution (provided in the kit) to each column.
Centrifuge the plate at 5,000 x g for 1 min at RT and discard the flow through.
Place the EZ-10 96-well plate back on the Deep Well Collection Plate and add 500 µL of universal NT solution (provided in the kit) to each column.
Centrifuge the plate at 5,000 x g for 1 min at RT and discard the flow through.
Place the EZ-10 96-well plate back on the Deep Well Collection Plate and centrifuge the column at 5,000 x g for 2 min at RT to ensure complete removal of the residual ethanol.
Place the EZ-10 96-well plate into a new deep well storage plate (provided in the kit) and add 30 µL RNase-free water (supplied in the kit), then incubate the plate at RT for 5 min.
Centrifuge the plate at 5,000 x g for 1 min at RT to elute the RNA solution.
Quantify the recovered RNA by measuring 1 µL of the RNA solution at 260 nm from about ten random wells using a NanoDrop. This will help estimate the isolated RNA concentrations.
Tightly seal the plate with an adhesive cover and keep it on ice (or -80 oC) until the RNA samples are ready for cDNA synthesis.
C. VSG-enriched cDNA synthesis and barcoding
This step requires a combination of primers (Fig 2) to barcode each clonal cell population cDNAs with a unique eight-nucleotide sequence for their identification during sequencing analysis. It will also provide an adapter sequence for DNA sequencing library preparation. The forward Ad-SL (5'-TTTCTGTTGGTGCTGATATTGCacagtttctgtactatattg-3') primer is universal and includes an Oxford nanopore adaptor sequence (capital letters) followed by a sequence (small letters) that hybridizes to mRNA splice leader sequence, a 39-nt sequence added to the 5' of all trypanosomes mRNAs. The reverse Ad-3endVSG primer (5'-TACTTGCCTGTCGCTCTATCTTCXXXXXXXXgtgttaaaatatatc-3') contains an Oxford nanopore adaptor sequence (capital letters) followed by eight nucleotide long variable sequence (barcode) unique to each clone, and a sequence pairing with the conserved 3'-end of VSG mRNAs, which encodes the C-terminus of VSG proteins (Mugnier et al., 2015). See Supplementary Information for the complete primer list. We recommend preparing a working primer solution containing a mix of both primers at 10 µM concentrations in a 96-well plate.
Take 4 μL of primer mix from the working primer solution plate and transfer it to each well of a new 96-well PCR plate using a multichannel pipette.
Add 1 μL of 10 mM dNTPs mix onto each well of the same 96-well PCR plate.
Thaw the RNA samples (from step B16) on ice if frozen at -80o C and add 5 μL of the samples keeping the same orientation in the 96-well plate as the original RNA plate. Mix the solutions by gently pipetting up and down, and spin down the plate briefly at 1,000 x g for 30 seconds.
Incubate the plate at 65 ºC for 5 min in a thermocycler, then transfer the plate immediately to ice.
Prepare cDNA synthesis master mix by adding 2 μl of MuLV 10x Buffer, 5U of M-MuLV Reverse Transcriptase, 8U of RNAse inhibitor (supplied in the MuLV reverse transcriptase kit) and adjust the reaction volume to 10 μl per reaction using Nuclease-free water.
Aliquot 10 μL of the cDNA synthesis master mix onto each well of the 96-well plate containing the RNA, dNTPs, and primer mix.
Thoroughly seal the plate using an adhesive plate sealing film, spin down at 1,000 x g for 30 seconds, and incubate at 42 ºC for 2 h and then 65 ºC for 20 min in a thermocycler.
Store the synthesized cDNA samples at -80 ºC or proceed to library preparation.
D. Oxford nanopore library preparation and DNA sequencing
Combine the synthesized cDNA samples from each well (from step C8) into one 1.5 mL microcentrifuge tube and mix gently. Avoid forming bubbles.
Prepare ten barcoding PCR reactions containing 5 μL of 10x Thermopol Buffer, 400 μM dNTPs mix, 500 nM barcode primer mix, 2U of Taq DNA Polymerase, 750 nM MgCl2. Add 3 μL of the pooled cDNA and adjust the final reaction volume to 50 μL with nuclease-free water.
Perform PCR reaction in a thermocycler at 95 ºC for 10 min, then 22 cycles of 95 ºC for 1 min, 62 ºC for 1 min and 68 ºC for 3:30 min, and a final extension at 68 ºC for 10 min. (See Notes 3 and 4)
Pool together all PCR amplicons in one 1.5 µL Eppendorf tube to obtain a final volume of 500 μL. Add 350 μL (0.65x beads to sample ratio) of NucleoMag NGS Clean-up and Size Selection beads to clean up the DNA.
Incubate beads/samples in a rotator (60 rpm) for 10 min at RT.
Spin down the tube briefly at 1,000 x g for 30 seconds, place it on a magnetic rack, and incubate for one min at RT. Collect and discard the supernatant.
Add 350 µL of freshly prepared 80% ethanol to the beads gently. Avoid disturbing the beads or taking the tube out of the magnetic rack. The volume of the 80% ethanol to add per wash should be equal to the sum of the total PCR amplicon volume and added NucleoMag NGS beads volume (e.g., 500 µL of PCR reaction + 350 µL of NucleoMag NGS magnetic beads = 850 µL of the mix, then 850 µL of 80% ethanol).
Incubate the mix for one min in the magnetic rack at RT. Collect and discard the 80% ethanol. Repeat steps 6 and 7.
Centrifuge the tube at 1,000 x g for 30 seconds. Place the tube back on the magnetic rack and remove any residual 80% ethanol from the tube.
Remove the tube from the magnetic rack and let beads air dry for 10 min at RT.
Resuspend beads in 61 μL nuclease-free water, mix gently by flicking the tube or gently pipetting up and down, and incubate for 5 min at RT.
Place the tube back on the magnetic rack and incubate for one minute at RT.
Collect and transfer the clear supernatant containing the eluted DNA into a new labelled tube. Discard the tube containing the beads.
Measure and assess the concentration and purity of the eluted DNA by quantifying 1 µL of the DNA sample using NanoDrop. The DNA yield ranges from about 30-60 ng/μL with 260/280 and 260/230 absorbance ratios of about 1.8 and 2.0, respectively.
Prepare 1 μg of DNA in 47 μL of nuclease-free water in a 0.2 mL thin-walled PCR tube, then add the following: 3.5 μL of NEBNext FFPE DNA Repair Buffer, 2 μL of NEBNext FFPE DNA Repair Mix, 3.5 μL Ultra II End-prep reaction buffer, 3 µL Ultra II End-prep enzyme mix, and 1 μL of DNA CS (DNA CS is provided in the Oxford Nanopore Ligation sequencing kit).
Mix the solutions by pipetting up and down gently, and incubate for 60 min at 20 °C in a thermocycler.
Add 39 μL of NucleoMag NGS Clean-up and Size Selection beads to the end-repaired DNAs (0.65x beads to samples ratio) to clean up and follow steps D5 to D13. Elute the DNA in 61 μL of Nuclease-free water.
Quantify 1 µL of the eluted DNA using NanoDrop. It is expected to recover about 90% of the DNA.
Transfer the remaining 60 µL end-repaired DNA volume from step 18 (0.5–1 μg) to a 0.2 mL thin-walled PCR tube, then add the following components: 25 μL of ligation buffer (LNB), 10 μL of NEBNext Quick T4 DNA ligase, and 5 μL of Adapter mix F (AMX-F) (LNB and AMX-F are components of the Oxford Nanopore Ligation sequencing kit). Mix by gently pipetting the reaction up and down, then briefly spin it down.
Incubate the reaction for 2 h at 20 °C in a thermocycler.
Transfer the ligation reaction mix from D20 to a new 1.5 mL Eppendorf tube and add 40 μL of well-resuspended AMPure XP beads to the reaction (0.4X beads to sample ratio). Mix by gently pipetting up and down.
Incubate the mix for 10 min rotating at RT.
Briefly centrifuge the sample at 1,000 x g for 30 seconds and place the tube on a magnetic stand for one min at RT or until the supernatant is clear and colourless.
Remove and discard the supernatant.
Resuspend the beads in 250 μL of Short Fragment Buffer (SFB), and mix gently by flicking the tube to wash the beads.
Briefly centrifuge the tube at 1,000 x g for 30 seconds. Transfer the tube to a magnetic stand to pellet the beads. Remove and discard the supernatant.
Repeat steps D25 and D26.
Briefly centrifuge the tube at 1,000 x g for 30 seconds. Place it on the magnetic stand and remove any residual supernatant. Air dry the beads at RT, and ensure that the beads do not dry to the point of cracking.
Remove the tube from the magnetic stand and resuspend the beads in 12 μL of Oxford nanopore Elution Buffer (EB) by gently pipetting up and down.
Briefly centrifuge the tube at 1,000 x g for 30 seconds and incubate for 10 min at RT.
Place the tube back onto the magnetic stand for one min, then remove and transfer the eluate containing the DNA library into a new 1.5 mL Eppendorf tube. Dispose of the beads.
Quantify 1 µL of the eluted library DNA sample using a NanoDrop. Expect a DNA library yield of approximately 10–40 ng/μL, (i.e., 200–1,500 fmol) of DNA.
Load 5–55 fmol of the prepared library into the flow cell (R9.4.1) and sequence the DNA according to the manufacturer's instructions.
Data analysis
A. Computational analysis of the sequenced library
The data analysis described here was performed using a Linux operating system (Ubuntu). The computational resources required will vary depending on the data available for analysis. The Oxford nanopore sequencer will generate fast5 files, which are basecalled to fastq files. The fastq files are the input dataset in the analysis. A standard fastq dataset with 100,906 reads was analyzed using 10 threads and 4 gigabytes of memory and completed in 5 minutes.
We developed a computational pipeline for the sequencing analysis and detection of VSG switching (Fig 2). The pipeline is run via the vsg-barseq.sh script. The pipeline takes the DNA sequences generated by the nanopore sequencing in fastq format and splits them into subfiles according to the cell barcodes (eight mers) used for DNA-seq library preparation. The output fastq file is named by its barcode, e.g., CCATGCAT.fastq. A split summary file (.txt) is generated and shows the number of reads per barcode file, the total count of reads analyzed, and the total amount of reads containing barcodes (see Note 5). Sequences from each file are then mapped to the organism genome (here T. brucei 427 strain) using minimap2, outputting .sam files. The alignments are then filtered using Samtools to remove supplementary alignments and to keep alignments with a mapQ ≥ 10. The resultant .sam files are sorted and indexed with Samtools resulting in sorted.bam and bam.bai files, respectively.
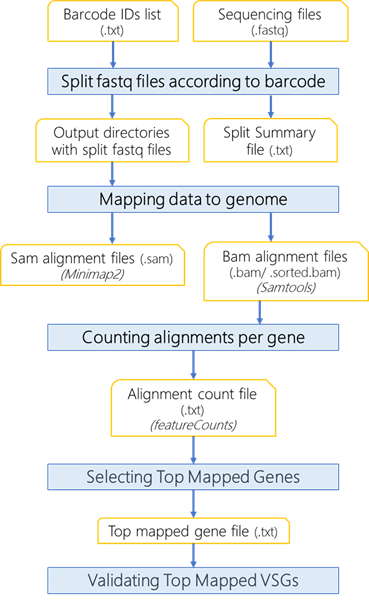
Figure 2. Flow chart of computational analysis using VSG-BarSeq. Multiplexed reads from clonal VSG-seq are split into clone-specific reads based on eight mers barcode. Reads are aligned to the genome using minimap2 and filtered with Samtools to remove supplementary and secondary alignments and keep alignments with mapQ ≥ 10. Then it counts the alignments per gene and reports the top alignment per file, corresponding to the expressed VSG gene.
The alignments are counted with featureCounts (package Subread). The top mapped reads, which correspond to the expressed VSG, are selected and compiled in a single tab-delimited file (topmapped.txt) while keeping the original output from featureCounts, which serves for analysis of other genes identified during alignment. The analysis of other identified genes is part of the quality control process. It typically shows genes with low counts reflecting low background noise from library preparation resulting primarily from sequences derived from splice-leader cDNA synthesis (Fig 3E).
The vsg-barseq.sh is open code and available on GitHub (https://github.com/cestari-lab/VSG-Bar-seq). The script takes six arguments:
1) directory of all files (output folders will be created in this directory)
2) directory of fastq files
3) directory of barcode.txt files (barcode.txt file has one barcode sequence per row)
4) directory of genome file in .fasta format
5) directory of gene transfer format file, i.e., .gtf format
6) number of threads to be passed to minimap2, samtools, and featureCounts (we recommend eight or more)
Example of barcode.txt file. Keep one barcode sequence per row.
CCGTTAGG
CCAACTAG
CAGGGCAG
CGCAGAAG
If the barcoded.txt file is generated using Windows operational system, we recommend converting from dos to Unix format before executing the analysis. The tools minimap2, samtools, subread, and rcgrep are required to run the script. After installing and/or loading the required tools, run the script as indicated below:
sh vsg-barseq.sh path/to/directory path/to/fastq path/to/barcodes path/to/genome path/to/gtf nthreads
See the example below using the structure folder exemplified (directories in bold).
sh vsg-barseq.sh \
c/user/vsg-barseq \
myfastq \
barcodes \
genomefiles/genome.fasta \
genomefiles/genome.gtf \
8
Directory structure of input files required to run the script:
c/user/vsg-barseq/
----myfastq/mydata.fastq
----barcodes/barcode.txt
----genomefiles/genome.fasta, genome.gtf
After running the script, the following output directories and files are created:
c/user/vsg-barseq/
----result_split_fastq/CCGTTAGG.fastq, CCAACTAG.fastq, CAGGGCAG.fastq, CGCAGAAG.fastq, split_summary.txt
----result_mapcount/
----sam/CCGTTAGG.sam, CCAACTAG.sam, CAGGGCAG.sam, CGCAGAAG.sam,
----bam/CCGTTAGG.bam, CCAACTAG.bam, CAGGGCAG.bam, CGCAGAAG.bam
----sorted_bam/CCGTTAGG_sorted.bam, CCGTTAGG_sorted.bam.bai, etc.
----counts/CCGTTAGG.txt, CCGTTAGG.txt.summary, etc.
----topmap/topmapped.txt
The result file topmapped.txt will have information on gene id (first row, in bold below), chromosome (or contig), nucleotide position (start, end), strand (+/-), gene length in nucleotides, number of counted alignments for the gene (seventh row, in bold below), and path to the original file containing all alignment counts for the corresponding clone; the eight mers barcode identifies the cell expressing the VSG. Each row represents a cell clone, and the gene id indicates the VSG expressed by the cells in the clonal population.
Tb427_000016000 BES1_Tb427v10 75570 77000 + 1431 461 /path/to/counts/CCGTTAGG.txt
Tb427_000016000 BES1_Tb427v10 75570 77000 + 1431 363 /path/to/counts/CCAACTAG.txt
Tb427_000008000 BES12_Tb427v10 45278 46651 + 1374 310 /path/to/counts/CAGGGCAG.txt
Tb427_000284800 Chr2_5A_Tb427v10 197226 198701 + 1476 840 /path/to/counts/CGCAGAAG.txt
B. Anticipated results
The analysis of parasite VSG switching starts with treating the cultures to induce the switching of the VSG gene. The treatment conditions and outcomes might differ for different cell lines, so they may need to be optimized. Here, the treatment was the temporal knockdown (24 h) of the PIP5Pase gene, which results in high rates of VSG switching (Touray et al., 2023). VSG switching occurs by alternating transcriptional between ESs or recombining VSG genes within ESs (Fig 3A). The parasite culture was diluted to obtain about 30% of clones from a 96-well plate. We recommend optimizing the dilution of the cells after the treatment, and checking cell viability, before the VSG-BarSeq experiment. After cDNA synthesis and library amplification with ONT-barcoding primers, we recommend analysis of the product in agarose gel. We often obtain a PCR fragment smear ranging from 200 bp to 2000 bp (Fig 3B).
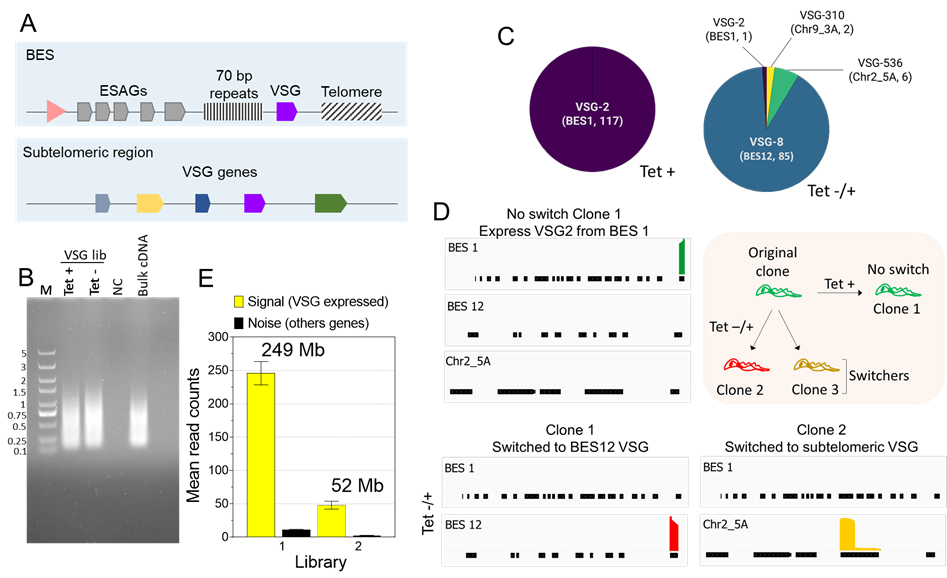
Figure 3. Analysis of VSG switching after PIP5Pase knockdown using VSG-BarSeq. (A) Diagram of bloodstream-form ESs (BES) and subtelomeric regions containing VSG genes used for recombination. The pink arrowhead represents a BES promoter. VSG genes are transcribed from BESs only. (B) Thirty-five cycles quality-control PCR amplification of pooled clonal VSG barcoded cDNAs. PCR was amplified with ONT barcode primers. NC, negative control, M, 5 kb DNA ladder. (C) VSG genes expressed by clones of T. brucei without PIP5Pase knockdown (Tet +) and after temporary knockdown for 24h followed by PIP5Pase re-expression and cloning for 5-7 (Tet -/+). Annotated names and corresponding chromosomes or BES identify VSG genes. The number of clones analyzed is indicated in parentheses. (D) Read coverage plots of example clones from Tet + or Tet -/+ treatment groups. The diagram on the top right summarizes the experiment and results—all clones derived from an original clone expressing VSG2 (BES 1). Graphs show read coverage of expressed VSGs on the same scale. (E) Signal-to-noise ratios of two different VSG-BarSeq libraries differing in sequencing depth. Sequencing depth is shown above plot bars as the total reads by the mean read length. Mb, megabases.
After library preparation, we usually obtain a library concentration ranging between 10–40 ng/μL with 260/280 and 260/230 ratios ~1.8 and 2.0, respectively. Library concentrations below 5ng/µL with considerable deviations from the above 260/280 and 260/230 ratios usually results in inferior quality and low throughput sequencing. Analysis of the sequencing using the vsg-barseq.sh script typically results in 70%–99% genome mapping. Fig 3C and D show the results of VSG switching in T. brucei after the temporary knockdown of PIP5Pase. Analysis of 117 clones from the control (no knockdown) showed that none of the cells switched VSGs (Fig 3C). However, after 24h temporary knockdown, there were 93 switchers out of 94 clones analyzed. There was a preference for the cells to switch to VSG8 in the BES12, suggesting transcriptional switching. Moreover, switching to VSGs from subtelomeric regions (Chr2_5A and Chr9_3A) was also detected, indicating switching by recombination (Fig 3C-D). The analysis does not require a significant amount of RNAs or sequencing throughput since the experimental setup relies on selecting VSG mRNAs, which are highly abundant (Cestari and Stuart, 2015), and on the analysis of clones rather than heterogeneous cell populations. Analysis of the expressed VSG (signal) compared to other genes (noise) showed a high signal-to-noise ratio, and increasing sequencing depth improved the signal without significantly increasing noise levels (Fig 3E). The amount of total RNA for cDNA synthesis per clone is minimal (5-20 ng), and sequencing depth required for analysis can be obtained from Oxford nanopore flongle flow cells, which typically produces 500-1,000 megabases of DNA sequencing (i.e., ~100,000 reads per group), thus reducing experimental costs. The results show the method's utility in identifying VSG switching events using multiplexed clonal cell barcodes. We anticipate that, with minimal modifications of the primers used for the target sequences, the approach can be applied to other gene families or other cell types, including var genes in Plasmodium sp., variant surface proteins in Giardia sp., as well as mucins and mucin-associated proteins (MASPs) in Trypanosoma cruzi. Analogously, the approach can be extended to other organisms or cell lines, including mammalian cells, e.g., T-cell or B-cell repertoire analysis.
General notes and troubleshooting
General notes
If more than 40% of the wells in a 96-well plate are positive, the parasites might not be clonal. We recommend optimizing the dilution of the parasites to obtain about a third of the wells from a 96-well plate containing growing parasites.
We recommend performing a quality control PCR prior cDNA amplification. Using conditions indicated in D3, but for 35 cycles. Then, analyze 30 µL the amplicons on 1% agarose/TAE electrophoresis gel with 5 µL of Ecostain at 85 Volts, for 45 min, and visualize using a gel imager. A smear migrating at 200 – 2,000 bp should be expected in the agarose gel (Fig 3B).
We recommend the volume of the pooled cDNA sample added to the PCR reaction be less than one-tenth of the total PCR reaction volume.
Reverse transcriptase enzymes are known to inhibit PCR, particularly at low template concentrations (Chandler et al., 1998). Therefore, adding more cDNA to the PCR reactions usually results in none or very little amplification.
The split summary file helps identify the splitting of reads into multiple barcode fastq files. If the number of reads with barcodes is larger than the total number of reads, it is indicative that some reads are included in more than one barcode fastq file. Primers with longer barcodes than eight mers could be used.
Table S1 – List of primer sequences indicated in the VSG-BarSeq protocol.
Acknowledgments
This research was funded by the Canadian Institutes of Health Research (grant CIHR PJT-175222 to IC); the Natural Sciences and Engineering Research Council of Canada (grant RGPIN-2019-05271 to IC); the Fonds de Recherche du Québec - Nature et technologie (grant 2021-NC-288072 to IC); and the Canada Foundation for Innovation (grant JELF 258389 to IC). AOT is a recipient of the Islamic Development Bank Scholarship (grant 600042744). This research was enabled in part by computational resources provided by Calcul Quebec (https://www.calculquebec.ca/en/) and the Digital Research Alliance of Canada (alliancecan.ca). This protocol is derived from the original research paper by Touray et al. (2023).
Competing interests
The authors declare that they have no conflicts of interest with the contents of this article.
References
Aitcheson, N., Talbot, S., Shapiro, J., Hughes, K., Adkin, C., Butt, T., Sheader, K. and Rudenko, G. (2005). VSG switching in Trypanosoma brucei: antigenic variation analysed using RNAi in the absence of immune selection. Mol Microbiol 57(6): 1608-1622.
Boothroyd, C. E., Dreesen, O., Leonova, T., Ly, K. I., Figueiredo, L. M., Cross, G. A. and Papavasiliou, F. N. (2009). A yeast-endonuclease-generated DNA break induces antigenic switching in Trypanosoma brucei. Nature 459(7244): 278-281. https://www.ncbi.nlm.nih.gov/pubmed/19369939
Cestari, I., McLeland-Wieser, H. and Stuart, K. (2019). Nuclear Phosphatidylinositol 5-Phosphatase Is Essential for Allelic Exclusion of Variant Surface Glycoprotein Genes in Trypanosomes. Mol Cell Biol 39(3).
Cestari, I. and Stuart, K. (2015). Inositol phosphate pathway controls transcription of telomeric expression sites in trypanosomes. Proc Natl Acad Sci U S A 112(21): E2803-2812.
Cestari, I. and Stuart, K. (2018). Transcriptional Regulation of Telomeric Expression Sites and Antigenic Variation in Trypanosomes. Curr Genomics 19(2): 119-132.
Chandler, D. P., Wagnon, C. A. and Bolton, H., Jr. (1998). Reverse transcriptase (RT) inhibition of PCR at low concentrations of template and its implications for quantitative RT-PCR. Appl Environ Microbiol 64(2): 669-677.
Danecek, P., Bonfield, J. K., Liddle, J., Marshall, J., Ohan, V., Pollard, M. O., Whitwham, A., Keane, T., McCarthy, S. A., Davies, R. M. and Li, H. (2021). Twelve years of SAMtools and BCFtools. Gigascience 10(2).
Faria, J., Glover, L., Hutchinson, S., Boehm, C., Field, M. C. and Horn, D. (2019). Monoallelic expression and epigenetic inheritance sustained by a Trypanosoma brucei variant surface glycoprotein exclusion complex. Nat Commun 10(1): 3023. https://www.ncbi.nlm.nih.gov/pubmed/31289266
Glover, L., Alsford, S. and Horn, D. (2013). DNA break site at fragile subtelomeres determines probability and mechanism of antigenic variation in African trypanosomes. PLoS Pathog 9(3): e1003260. https://www.ncbi.nlm.nih.gov/pubmed/23555264
Jehi, S. E., Li, X., Sandhu, R., Ye, F., Benmerzouga, I., Zhang, M., Zhao, Y. and Li, B. (2014). Suppression of subtelomeric VSG switching by Trypanosoma brucei TRF requires its TTAGGG repeat-binding activity. Nucleic Acids Res 42(20): 12899-12911.
Li, H. (2018). Minimap2: pairwise alignment for nucleotide sequences. Bioinformatics 34(18): 3094-3100.
Liao, Y., Smyth, G. K. and Shi, W. (2014). featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30(7): 923-930.
Lopez-Escobar, L., Hanisch, B., Halliday, C., Ishii, M., Akiyoshi, B., Dean, S., Sunter, J. D., Wheeler, R. J. and Gull, K. (2022). Stage-specific transcription activator ESB1 regulates monoallelic antigen expression in Trypanosoma brucei. Nat Microbiol 7(8): 1280-1290. https://www.ncbi.nlm.nih.gov/pubmed/35879525
Mugnier, M. R., Cross, G. A. and Papavasiliou, F. N. (2015). The in vivo dynamics of antigenic variation in Trypanosoma brucei. Science 347(6229): 1470-1473.
Ramirez, F., Ryan, D. P., Gruning, B., Bhardwaj, V., Kilpert, F., Richter, A. S., Heyne, S., Dundar, F. and Manke, T. (2016). deepTools2: a next generation web server for deep-sequencing data analysis. Nucleic Acids Res 44(W1): W160-165.
Rudenko, G., Chaves, I., Dirks-Mulder, A. and Borst, P. (1998). Selection for activation of a new variant surface glycoprotein gene expression site in Trypanosoma brucei can result in deletion of the old one. Mol Biochem Parasitol 95(1): 97-109.
Touray, A. O., Rajesh, R., Isebe, T., Sternlieb, T., Loock, M., Kutova, O. and Cestari, I. (2023). A PI(3,4,5)P3-dependent allosteric switch controls antigenic variation in trypanosomes. eLife 12: RP89331.
Ulbert, S., Chaves, I. and Borst, P. (2002). Expression site activation in Trypanosoma brucei with three marked variant surface glycoprotein gene expression sites. Mol Biochem Parasitol 120(2): 225-235.
Supplementary information
Table S1 – List of primer sequences indicated in the VSG-BarSeq protocol.
Related files
 Cestari_manuscript.docx Supplementary Information.docx
Cestari_manuscript.docx Supplementary Information.docx - Touray, A O, Sternlieb, T, Isebe, T and Cestari, I(2023). Identifying antigenic switching by clonal cell barcoding and nanopore sequencing in trypanosomes. Bio-protocol Preprint. bio-protocol.org/prep2385.
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.