

Myonecrosis Induction by Intramuscular Injection of CTX
肌内注射 CTX 诱导肌肉坏死
(*contributed equally to this work) 发布: 2023年01月05日第13卷第1期 DOI: 10.21769/BioProtoc.4587 浏览次数: 3975
评审: Vivien Jane Coulson-ThomasDieu-Huong HoangMarielle Saclier
参见作者原研究论文
The authors used this protocol in:

Nov 2021
Advertisement








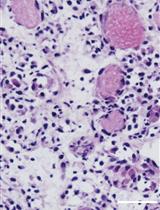
Abstract
Skeletal muscle, one of the most abundant tissue in the body, is a highly regenerative tissue. Indeed, compared to other tissues that are not able to regenerate after injury, skeletal muscle can fully regenerate upon mechanically, chemically, and infection-induced trauma. Several injury models have been developed to thoroughly investigate the physiological mechanisms regulating skeletal muscle regeneration. This protocol describes how to induce muscle regeneration by taking advantage of a cardiotoxin (CTX)-induced muscle injury model. The overall steps include CTX injection of tibialis anterior (TA) muscles of BL6N mice, collection of regenerating muscles at different time points after CTX injury, and histological characterization of regenerating muscles. Our protocol, compared with others such as those for freeze-induced injury models, avoids laceration or infections of the muscles since it involves neither surgery nor suture. In addition, our protocol is highly reproducible, since it causes homogenous myonecrosis of the whole muscle, and further reduces animal pain and stress.
Graphical abstract

Background
The final goal of regenerative medicine is to reconstitute tissue and organ functionality after injury or disease (Mao and Mooney, 2015). Tissue regeneration is a physiological process orchestrated by different infiltrating and tissue-resident cell types. Skeletal muscle is one of the most dynamic and plastic tissues of the human body and represents approximately 40% of total body weight in humans (Frontera and Ochala, 2015). Skeletal muscle not only controls posture maintenance and locomotion but plays a fundamental role in the maintenance of metabolic homeostasis, being involved in heat production and carbohydrate and amino acid storage (Schiaffino et al., 2013). In this context, muscle degeneration due to acute and chronic conditions leads to reduced mobility and strength, and to metabolic disorders (Schiaffino et al., 2013). Regeneration of skeletal muscle relies on a tightly regulated gene expression program and on the activation of specific signaling pathways that characterize muscle embryonic development (Chargé and Rudnicki, 2004). The ability of muscle to regenerate is principally due to a specific population of normally quiescent muscle stem cells called satellite cells that are strictly associated with muscle fibers (Chargé and Rudnicki, 2004). Satellite cells are myogenic precursor cells (MPCs) that reside in a quiescent state beneath the basal lamina that surrounds muscle fibers (Chargé and Rudnicki, 2004). In detail, soon after muscle damage, satellite cells escape quiescence and start to proliferate. A subset of these activated cells proliferate and differentiate, whereas others return to quiescence to reconstitute the pool of satellite cells that will be crucial for further rounds of skeletal muscle regeneration (Chargé and Rudnicki, 2004).
Although satellite cells play a key role in muscle regeneration, their presence is not sufficient for efficient skeletal muscle repair, and many additional cell types play an active role in promoting tissue repair (Joe et al., 2010; Uezumi et al., 2010). Among them, key players in the complex scenario of skeletal muscle regeneration are inflammatory cells that invade muscle soon after injury and, together with satellite cells, contribute to a complete muscle regeneration (Tidball, 2017). Among the inflammatory cells, monocytes/macrophages play a major role in the repair process (Wang and Zhou, 2022). For example, macrophages release cytokines and growth factors that contribute to satellite cell activation, proliferation, and differentiation, according to their polarization state (Saclier et al., 2013). In particular, soon after muscle damage, macrophages adopt a pro-inflammatory profile, releasing pro-inflammatory cytokines such as tumor necrosis factor alpha (TNF-α), interleukine-6 (IL-6) and insulin-like growth factor (IGF-1) that, in turn, promote satellite cell proliferation (Tidball, 2017). Starting from three to five days post injury, these macrophages skew from a pro- to an anti-inflammatory profile, releasing anti-inflammatory cytokines that not only dampen the inflammatory response but also play trophic function by promoting satellite cell differentiation and fusion, thus contributing to the formation of new myofibers (Tidball, 2017). To date, different mouse models of acute muscle injury are available, as they represent an attractive system for investigating interactions between the immune system and satellite cells during skeletal muscle regeneration. Indeed, the onset of tissue damage is well defined and the time course of inflammation and regeneration predictable (Tidball, 2017). The most used injury models are freeze injury (FI), barium chloride injection, notexin (NTX) injection, and cardiotoxin (CTX) injection. The last one provides a useful model for sterile inflammation and, importantly, induces homogeneous damage to the whole muscle. Furthermore, it triggers the infiltration of many monocytes and macrophages participating in muscle repair, regeneration, and growth (Rigamonti et al., 2014). Here, we describe a protocol to induce muscle sterile injury by CTX injection. This method allowed us to study the influence of mitochondrial calcium uptake on macrophage metabolic profiles and on the skeletal muscle regeneration process (Feno et al., 2021).
Myonecrosis is induced by intramuscular injection of CTX in tibialis anterior (TA) muscles, as performed in many other laboratories (Mounier et al., 2013; Perdiguero et al., 2011; Saclier et al., 2013). Compared with other protocols, our method has the great advantage of avoiding muscle laceration (Feno et al., 2021). Specifically, we can directly inject CTX in TA muscles without exposing them, thus avoiding surgery and consequent damage. Muscles are collected for analysis at different time points after injury (at 3, 7, and 14 days) to precisely characterize the time course of skeletal muscle regeneration. After muscle collection, regenerating TA muscles are frozen, and muscle sections are characterized by hematoxylin and eosin (H&E) staining to evaluate the efficiency of the CTX- induced skeletal muscle regeneration process.
Materials and Reagents
8 weeks C57BL6N male mice (Charles River)
Isoflurane 1000 mg/g (Piramal Critical Care, catalog number: 803249)
500 μM CTX (Merk, catalog number: 217503-1MG)
9% NaCl2 (Merk, catalog number: S3014)
UltraPure DNase/RNase-Free distilled water (Thermo Fisher Scientific, catalog number: 10977049)
70% v/v ethanol (Merk, catalog number: 64-17-5)
Isopentane (2-Methylbutane anhydrous ≥99%-1l) (Merck, catalog number: 277258)
Liquid nitrogen
Hematoxylin & Eosin (H&E) kit staining (Bio-Optica, catalog number: 04-061010)
Optimal cutting temperature compound (O.C.T.) (Tissue-Tek, catalog number: 4583)
Equipment
VetFlo veterinary anesthesia apparatus
Dissection tools:
Fine scissors (Fine Science Tools, catalog number: 14160-10)
Fine forceps (Fine Science Tools, catalog number: 11412-11)
Standard forceps (Fine Science Tools, catalog number: 11150-10)
Microcentrifuge tubes 1.5 mL (Sarstedt, catalog number: 72706)
Cryovials (Sarstedt, catalog number: 72.694.006)
Glass slides (Vetrotecnica, catalog number: 01.4230.34)
500 μL syringe 30 G (Vetrotecnica, catalog number: 11.3525.45)
Plastic scoop
Dewar bottle for liquid nitrogen
Cryostat (Leica, model: CM1850)
Procedure
文章信息
版权信息
© 2023 The Authors; exclusive licensee Bio-protocol LLC.
如何引用
Readers should cite both the Bio-protocol article and the original research article where this protocol was used:
- Feno, S., Munari, F., Gherardi, G., Vecellio Reane, D., D’Angelo, D., Viola, A., Rizzuto, R. and Raffaello, A. (2023). Myonecrosis Induction by Intramuscular Injection of CTX. Bio-protocol 13(1): e4587. DOI: 10.21769/BioProtoc.4587.
- Feno, S., Munari, F., Reane, D. V., Gissi, R., Hoang, D. H., Castegna, A., Chazaud, B., Viola, A., Rizzuto, R. and Raffaello, A. (2021). The dominant-negative mitochondrial calcium uniporter subunit MCUb drives macrophage polarization during skeletal muscle regeneration. Sci Signal 14(707): eabf3838.
分类
细胞生物学 > 组织分析 > 损伤模型
细胞生物学 > 细胞成像 > 冷冻超薄切片
您对这篇实验方法有问题吗?
在此处发布您的问题,我们将邀请本文作者来回答。同时,我们会将您的问题发布到Bio-protocol Exchange,以便寻求社区成员的帮助。