

Assay to Study the Phase-transition Behavior of Edc3, a Conserved Processing Body (P-body) Marker Protein
研究保守的加工体 (P-body) 标记蛋白Edc3 相变行为的试验
发布: 2022年08月20日第12卷第16期 DOI: 10.21769/BioProtoc.4487 浏览次数: 3455
评审: Julie WeidnerIndranil MalikAnu P. Minhas
参见作者原研究论文
The authors used this protocol in:

Apr 2022
Advertisement










Abstract
RNA granules are conserved, non-membranous, biphasic structures predominantly composed of RNA and RNA-binding proteins. RNA granules often assemble as a result of cellular responses to a variety of stresses, including infection. Two types of RNA granules are best characterized: stress granules (SGs) and processing bodies (P-bodies). The mechanism of RNA granule assembly and disassembly is still understudied because of its complex composition and dynamic behavior. The assembly of RNA granules is driven by a process known as phase separation of granule components. Edc3 is a conserved decapping activator and an essential P-body component in Saccharomyces cerevisiae. Phase separation of P-body proteins has been poorly explored. This protocol will enable the visualization of the phase transition behavior of Edc3, since it is tagged to mCherry. It further describes using small molecules and other proteins to study P-body dynamics. In addition to the assembly of Edc3, this assay also lays the foundation to study disassembly of phase-separated assemblies in vitro, which was not explored earlier. We have devised the assay to describe the use of one such protein that acts as a disassembly factor. Overall, this protocol is simple to perform and can potentially be combined with analyzing these assemblies using other approaches.
Graphical abstract:
Background
RNA granules are mRNA-protein complexes that function as sites of mRNA storage and/or degradation (Anderson and Kedersha, 2006). Cytoplasmic RNA granules are also known as higher-order messenger ribonucleoprotein (mRNP) complexes (Anderson and Kedersha, 2009). RNA granules are dynamic and can exchange mRNP components with their surroundings (Bhattacharyya et al., 2006; Wheeler et al., 2016). RNA granules are also insoluble in lysates, non-membranous cytoplasmic complexes, or foci containing translationally repressed and/or degrading mRNAs. They are found in various cells, including, but not limited to yeast, germ cells, embryos, and neurons, often in response to stress (Thomas et al., 2011; Wheeler et al., 2016). Upon release from RNA granules, the mRNAs can return back to translation. Thus, RNA granules could be mRNA fate-determining sites (Mitchell and Parker, 2014). RNA granules share certain similarities across species (Anderson and Kedersha, 2006), suggesting that they are conserved and share mechanisms of granule formation and regulation of mRNA fate (Zlotorynski, 2015). Stress granules and P-bodies are two important types of cytoplasmic RNA granules that are majorly studied in the field of RNA granule biology. Stress granules (SGs) assemble in the cytoplasm upon stress and are known to harbor translationally repressed mRNAs (Buchan et al., 2011; Rajyaguru et al., 2012). On the other hand, processing bodies (P-bodies) are visible even in the absence of stress but grow in size and intensity in response to stress (Buchan et al., 2011).
Previous reports suggested an important role of intrinsically disordered regions (IDRs) during the assembly of RNA granules (Figure 1A), which has been extensively studied (Jonas and Izaurralde, 2013; Protter et al., 2018). IDRs are specific amino acid sequences or repeats in a protein that do not fold into a secondary structure. The RGG-repeats (arginine-glycine-glycine repeats), a well-studied IDR, play an essential role in RNA granule assembly and translation repression (Rajyaguru et al., 2012; Poornima et al., 2016; Brandariz-Núñez et al., 2018; Ozdilek et al., 2017; Bhatter et al., 2019). Single-stranded nucleic acid-binding protein 1 (Sbp1) contains N-terminal RRM1, C-terminus RRM2, and an RGG-motif sandwiched between the RRM domains (Bhatter et al., 2019). Previous reports suggest the RGG-motif of Sbp1 is essential for binding to the translation initiation factor eIF4G and repressing translation (Rajyaguru et al., 2012; Bhatter et al., 2019). Sbp1 orchestrates P-body disassembly with the help of its RGG-motif (Roy et al., 2022) (Figure 1B).
Several recent reports have found that RNA binding proteins (RBPs), such as G3BP1, PABP1, Tau, FUS, TDP-43, α-synuclein, and hnRNPs, can undergo liquid-liquid phase separation (LLPS). LLPS is a process in which proteins and nucleic acids in cells transition from a liquid phase to a condensed state resembling a liquid droplet (Figure 1A). In neurodegeneration, mutations in RBPs led to aberrant cytoplasmic localization. This mislocalization could result in protein transition to a gel or solid phase (irreversible state) rather than the liquid condensate formation (reversible state) in the cell cytoplasm, promoting aggregation and fibril assembly (Couthouis et al., 2011; Lin et al., 2015; Wheeler et al., 2016). Persistent aggregation and fibril assembly of these RBPs in the cytoplasm can be toxic and lethal for cell survival, as reported in the context of certain neurodegenerative disorders, such as ALS and FTD (Sun et al., 2011; Brunello et al., 2016). Recently, α-synuclein has been found to modulate P-body dynamics by physically binding to decapping factors, disrupting P-body composition, and altering mRNA decay dynamics. Binding of α-synuclein to P-body components alters the dynamics of P-bodies, which act as feedback for α-synuclein aggregates in the cytoplasm (Hallacli et al., 2022).
There are few reports suggesting RBPs undergo phase separation, which is involved in the formation of RNA granules, such as stress granules (Mugler et al., 2016; Riback et al., 2017). Here, we report an extended assay that describes phase separation of Edc3, a conserved P-body marker protein. Unlike stress granules, proteins involved in decapping and degradation of mRNA are found to localize within the P-bodies, therefore believed to be the hub for mRNA decay (Buchan et al., 2011; Mitchell et al., 2013) (Figure 1A). Edc3 is a P-body resident protein and plays an important role in the formation and maintenance of P-bodies. Edc3 contains a C-terminal YjeF-N motif, which can self-interact, thereby contributing to P-body assembly (Decker et al., 2007) (Figure 1A). Deletion of Edc3 leads to poor P-body formation (Decker et al., 2007). Edc3 interacts with RPS28B mRNA, and this interaction has recently been reported to be important for efficient P-body formation (Fernandes and Buchan, 2020). To sum up, Edc3 self-association and interaction with P-body resident proteins and RNAs contribute to higher-order P-body assemblies. Like any conventional RNA granules, P-bodies are very dynamic and shuttle protein-RNA components in and out of the granules (Aizer et al., 2008). This dynamic nature of P-bodies makes them very difficult to study. Therefore, to study the dynamics of P-bodies, we need to develop an in vitro assay system, which must be in accordance with RNA granule studies and can easily be performed in the laboratory. In this protocol, we discuss a multi-component assay system that can be used to study P-body dynamics (assembly and disassembly) in vitro.

Figure 1. Illustrations depicting assembly and disassembly of processing bodies (P-bodies). (A) Self-association of Edc3, recruitment of other P-body resident proteins, along with the association of mRNA and co-factor like NADH, facilitates phase transition of Edc3 leading to P-body assembly. (B) During recovery, Sbp1, with the help of its RGG-motif (IDR), binds to the YjeF-N domain of Edc3, disrupting the self-association and leading to P-body disassembly.
Development of the protocol
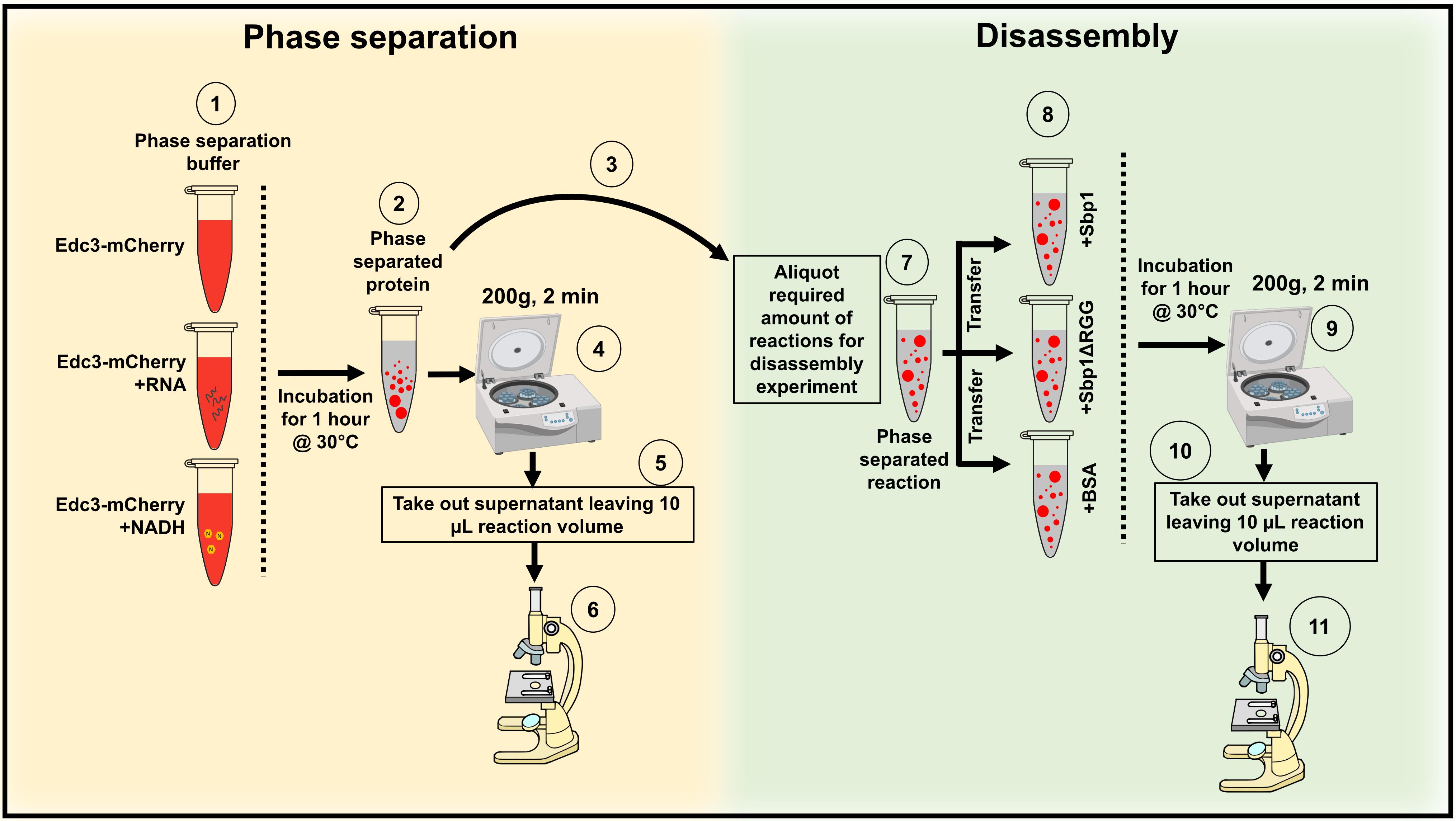
This protocol has been developed to understand the phase separation behavior of Edc3-mCherry in vitro (Roy et al., 2022) (Figure 2). Edc3 is a conserved core P-body protein, which is important for forming and maintaining P-bodies. The study of Edc3 assemblies in vitro is key to understanding the process of Edc3 phase separation in vivo and its regulation. Using this protocol, we observe that after 1 h of incubation, purified Edc3-mCherry can form small assemblies in phase separation buffer (PSB) (Figure 3D), which grow in size and intensity in the presence of RNA or NADH (Figure 3E–F), compared to 0 h (Figure 3A–C). We have extended this assay to analyze the impact of a purified disassembly factor on Edc3 assemblies. We found that adding purified Sbp1 to Edc3-mCherry assemblies leads to its dissociation (Figure 4A–B). Overall, this assay provided two important results: a) RNA and NADH promote the formation of Edc3 assemblies, and b) Sbp1 leads to the dissolution of Edc3 assemblies.
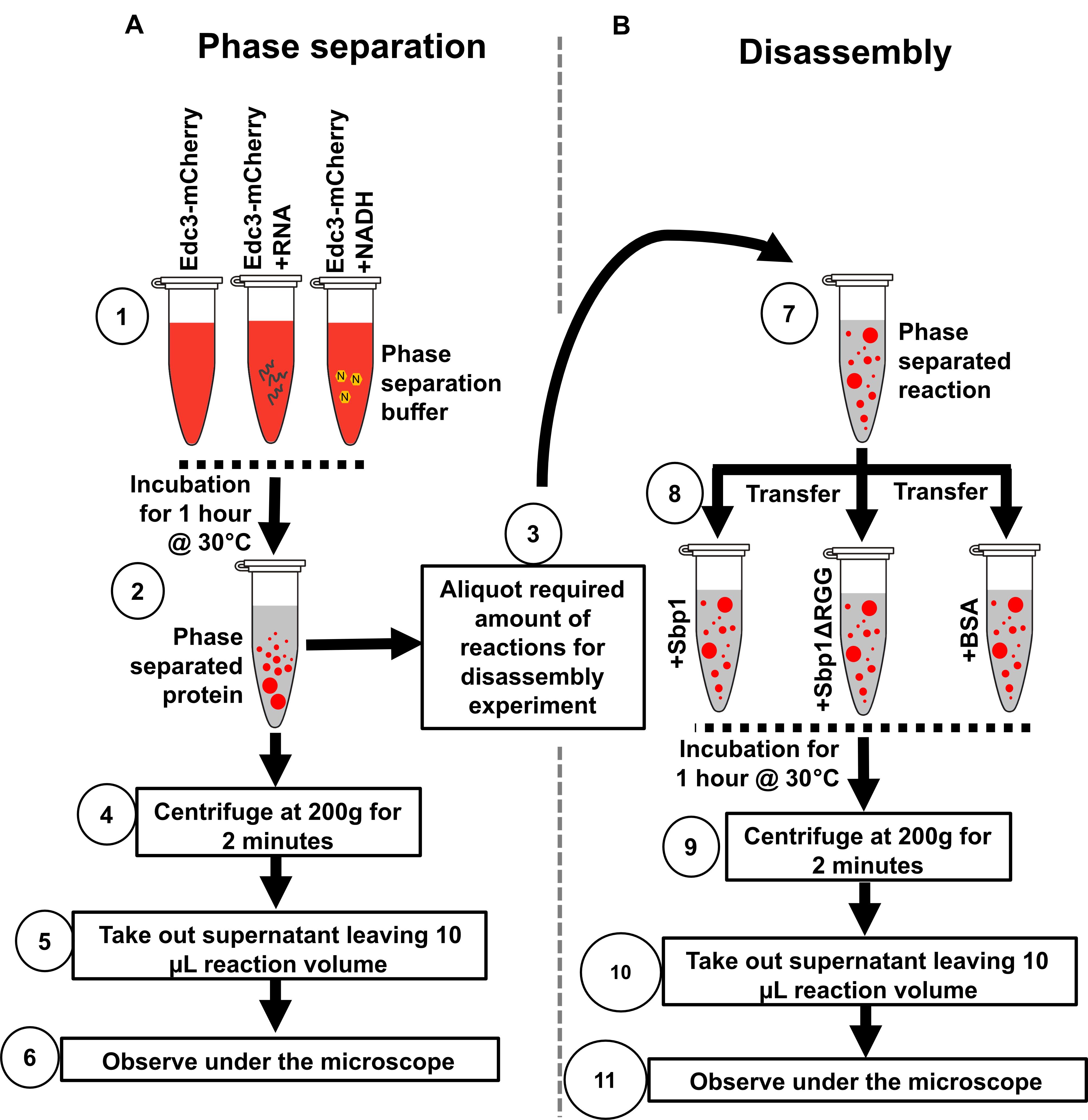
Figure 2. Illustration depicting the protocol steps to study the phase separation of Edc3 in vitro. (A) Phase separation behavior of Edc3. (B) Disassembly of phase-separated Edc3.
Applications of the method
This method can be optimized to perform high-throughput screening to identify small molecules that can perturb assembly and/or disassembly of P-bodies. Such small molecules could also be tested for their ability to perturb the aggregates of proteins implicated in neurodegenerative disorders. mRNAs with specific sequences and structures can be tested for their role in promoting Edc3 assembly or disassembly. Furthermore, purified proteins can be added to the assay system to identify factors affecting the assembly and/or disassembly of higher-order Edc3 structures.
Comparison with other methods
Most phase-separation studies use a buffer containing crowding reagents, like BSA, PEG, or glycerol (Lin et al., 2015; Mugler et al., 2016), to reduce the surface area of the reaction in the buffer system. We have standardized this assay such that it does not contain reagents that may directly or indirectly affect protein stability, structure, or function. Also, we wanted an efficient buffer condition for studying both the assembly and disassembly of Edc3 in vitro. We narrowed it down to KCl, HEPES-KOH, and MgCl2, suitable for the Edc3-mCherry assembly and disassembly experiments without compromising protein stability.
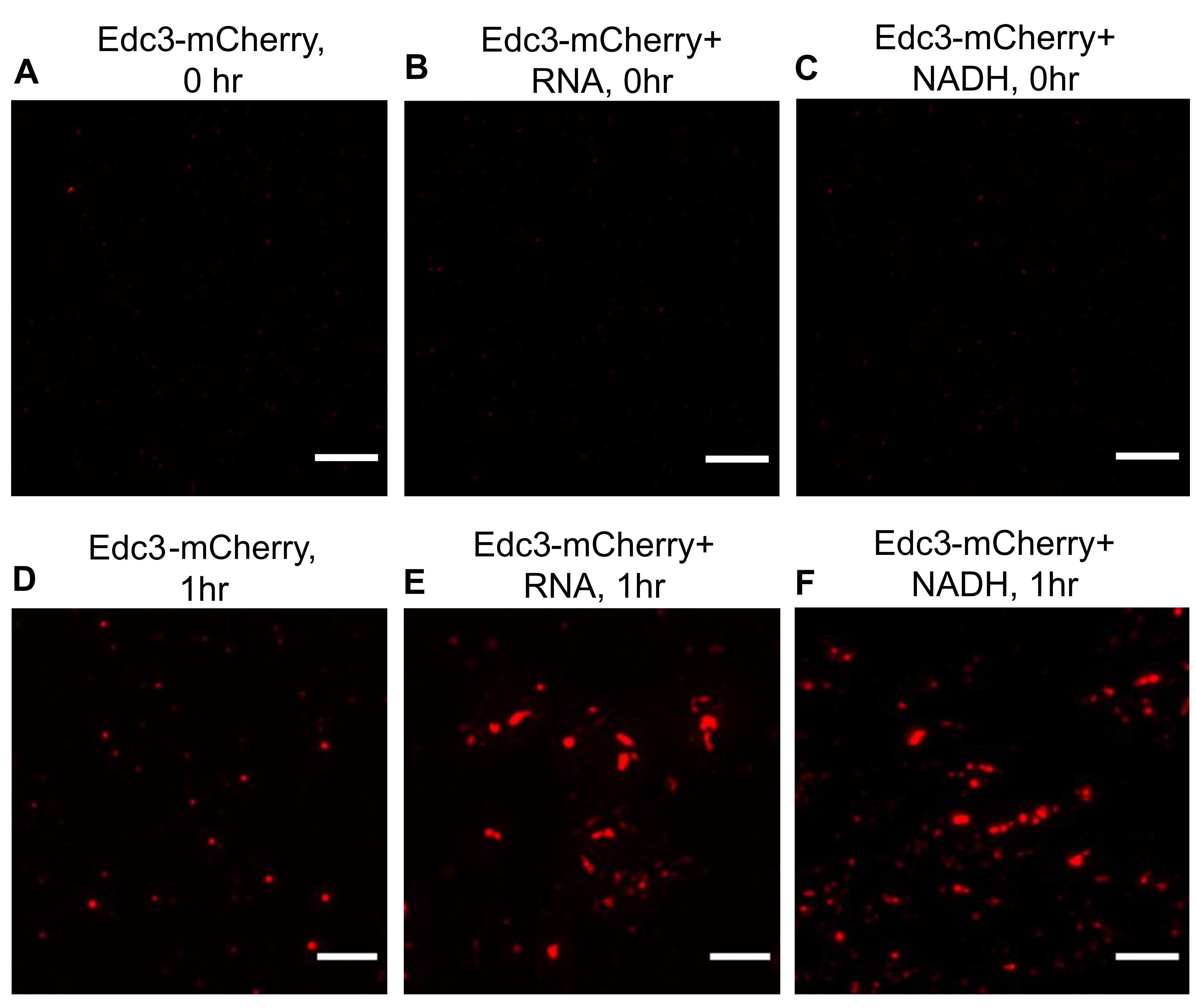
Figure 3. Phase separation of Edc3-mCherry in the presence of RNA and NADH. (A) Phase separation of Edc3-mCherry alone at 0 h. (B) Phase separation of Edc3-mCherry in the presence of RNA, (C) or NADH at 0 h. (D) Phase separation of Edc3-mCherry alone after 1 h. (E) Phase separation of Edc3-mCherry in the presence of RNA, (F) or NADH after 1 h at 30 °C. The scale bar represents 5 µm.
Expertise needed to implement the protocol
This protocol has been standardized so that it can be performed by any researcher with a basic knowledge of Biochemistry, protein chemistry, and wide-field fluorescence microscopy.
Limitations
RNA granules contain a variety of RNA binding proteins with different physical properties. Most of the granule residing proteins contain intrinsically disordered regions in different proportions. IDR plays a vital role in the phase separation of granule resident proteins. Therefore, it is unlikely that all IDR-containing proteins will phase-separate under similar conditions. This protocol is designed and standardized for phase separation of Edc3-mCherry in vitro. However, tweaking this protocol may also be useful for studying the phase separation of other RNA-binding proteins.
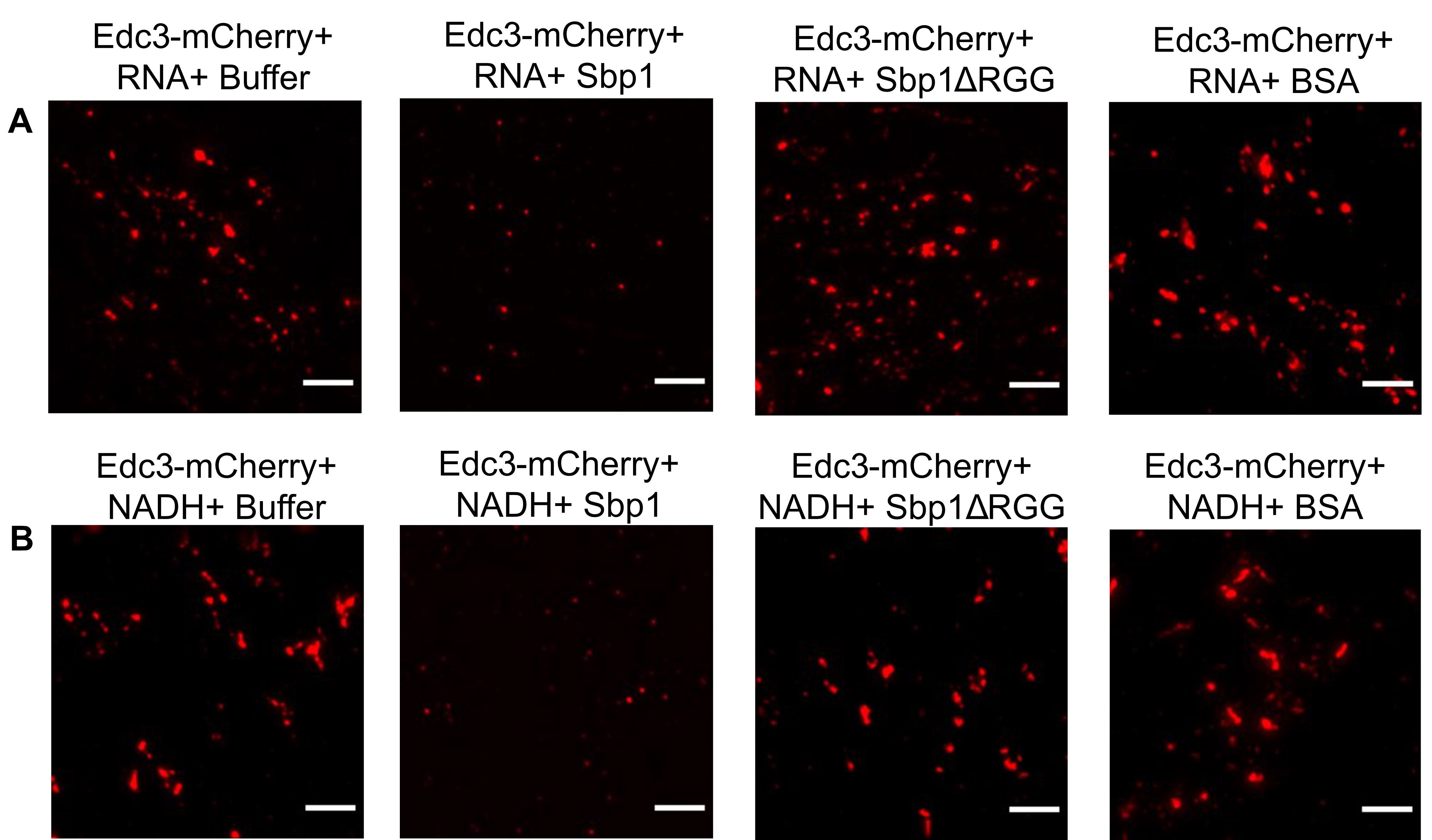
Figure 4. Disassembly of phase-separated Edc3. (A) Disassembly of phase-separated Edc3+RNA assemblies in the presence of either PSB, Sbp1, Sbp1ΔRGG, or BSA. (B) Disassembly of phase-separated Edc3+NADH assemblies in the presence of either PSB buffer, Sbp1, Sbp1ΔRGG, or BSA. The scale bar represents 5 µm.
Materials and Reagents
1.5 mL microcentrifuge tube (Axygen, catalog number: MCT-150-C)
Frosted microscope slides (Bluestar, 75 mm × 25 mm, 1.35 mm thickness)
Microscope cover glasses (Bluestar, Square 22 mm, 0.13 mm to 0.16 mm thickness)
Microcentrifuge tube holder (VWR, catalog number: 211-0207)
Purified proteins- Edc3-mCherry, Sbp1, Sbp1ΔRGG (Figure 5A) are stored at -80 °C with 10% glycerol in dialysis buffer (Roy et al., 2021). Purified proteins must be kept in aliquots at -20 °C (maximum three days as these proteins are prone to precipitation) or -80 °C (for long-term use, ≥ one week), DO NOT freeze-thaw the proteins more than two times; this might affect protein stability.
Total RNA isolated from Saccharomyces cerevisiae (Figure 5B) using the hot phenol method (store the RNA at -20 °C for short-term and -80 °C for long-term) (Garg et al., 2020).
Bovine Serum Albumin (BSA; Sigma, catalog number: A4503-100G) is stored at 4 °C.
Potassium chloride (KCl; Sigma, catalog number: P9541-1KG) is stored at room temperature.
N-(2-Hydroxyethyl) Piperazine N-(2-Ethane Sulphonic Acid) [HEPES buffer; SRL, catalog number: 16826] is stored at 4 °C.
Magnesium chloride (MgCl2; Sigma, catalog number: M0250-1KG) is stored at room temperature.
Potassium hydroxide (KOH; Sigma, catalog number: V800320-500G) is stored at room temperature.
Diethyl pyrocarbonate (DEPC; SRL, catalog number: 46791) is stored at 4 °C.

Figure 5. Purified proteins and total RNA used in this assay. (A) Purified Edc3-mCherry, only-mCherry, Sbp1, and Sbp1ΔRGG, used for the phase separation and disassembly experiments. * Represents a degradation product of Edc3-mCherry. (B) 0.8% Formamide agarose gel electrophoresis depicting the quality of total RNA isolated from S. cerevisiae cells. Bands represent 25S rRNA, 18S rRNA, and small RNA species, respectively.β-Nicotinamide adenine dinucleotide, reduced disodium salt hydrate (NADH; Sigma, catalog number: N8129-100MG) is stored at -20 °C.
Immersion oil 1.516 NA (Cargille, catalog number: 20130)
Lint-free tissue paper (Kimwipes, Kimtech, catalog number: 34155)
500 mM KCl (For reagent preparation, see Recipes) should be stored at 4 °C.
150 mM HEPES-KOH, pH 7.4 (For buffer preparation, see Recipes), should be stored at 4 °C.
100 mM MgCl2 (For reagent preparation, see Recipes), should be stored at 4 °C.
0.2 mM NADH (For reagent preparation, see Recipes), aliquot, and store at -80 °C. NADH is prone to degradation; therefore, it is important to aliquot 1mM NADH in nuclease-free water and store it at -80 °C.
DEPC treated H2O/Nuclease-free H2O (For reagent preparation, see Recipes), store at room temperature.
Phase Separation Buffer (PSB; For buffer preparation, see Recipes) must be prepared on ice. All the reaction steps must be done on ice (or 4 °C), and after reaction setup, transfer it to respective temperatures.
Absolute ethanol (EMSURE®, Merck, catalog number: 100983)
70% ethanol (see Recipes)
DEPC treated water (see Recipes)
500 mM Potassium Chloride (KCl) (see Recipes)
150 mM HEPES-KOH buffer, pH 7.4 (see Recipes)
100 mM Magnesium chloride (MgCl2) (see Recipes)
Phase separation buffer (PSB) (see Recipes)
Notes:
Store all the buffers at 4–8 °C. DO NOT store them at room temperature.
Wash the coverslips and glass slides with ethanol and dry them using lint-free tissue paper. Dust can hinder visualizing the phase-separated proteins.
Alternative to DEPC-treated water or solutions, commercially available RNase-free reagents can also be used for the assay.
70% ethanol is used to wash or clean the equipment. Alternatively, decontaminating agents like RNaseZap could also be used.
Equipment
Autoclavable pipettes (1–10 µL, 2–20 µL, 20–200 µL; Finpipette,Thermo Fisher Scientific, catalog number: 4701070). Clean the pipette using 70% ethanol before proceeding with the assay.
Autoclaved DNase, RNase, pyrogen-free micro tips (Axygen, catalog numbers: T-300; T-200-Y)
70% ethanol wiped scissors for cutting tip nose
30 °C incubator/heat block (Shalom, catalog number: SLM-INC-270)
Vortex Mixer (Neuation, catalog number: VM-2110)
Eppendorf® Centrifuge 5424R (Eppendorf, catalog number: EP5404000537)
DeltavisionTM Elite Microscope (GE Healthcare) with 100× objective
Software
softWoRx 6.1.3 software for image acquisition (Applied Precision, LLC)
Fiji ImageJ Version 1.53f51 for image processing (Open source, https://imagej.net/software/fiji/downloads)
Microsoft Excel (Microsoft Corporation, USA)
GraphPad Prism 4 for data analysis (GraphPad Software Inc., https://www.graphpad.com/)
Procedure
文章信息
版权信息
© 2022 The Authors; exclusive licensee Bio-protocol LLC.
如何引用
Roy, R. and Rajyaguru, P. I. (2022). Assay to Study the Phase-transition Behavior of Edc3, a Conserved Processing Body (P-body) Marker Protein. Bio-protocol 12(16): e4487. DOI: 10.21769/BioProtoc.4487.
分类
生物化学 > 蛋白质 > 分离和纯化
分子生物学 > RNA > RNA-蛋白质相互作用
您对这篇实验方法有问题吗?
在此处发布您的问题,我们将邀请本文作者来回答。同时,我们会将您的问题发布到Bio-protocol Exchange,以便寻求社区成员的帮助。