

Isolation and Culture of Cranial Neural Crest Cells from the First Branchial Arch of Mice
小鼠第一鳃弓颅神经嵴细胞的分离培养
发布: 2022年04月05日第12卷第7期 DOI: 10.21769/BioProtoc.4371 浏览次数: 3104
评审: Chiara AmbrogioJohn W PetersonMaría Delgado EstebanAnonymous reviewer(s)
参见作者原研究论文
The authors used this protocol in:

Jan 2021
Advertisement








Abstract
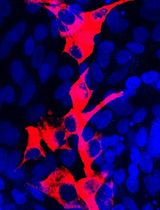
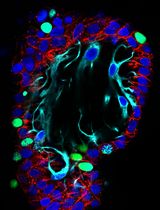
Craniofacial anomalies (CFA) are a diverse group of deformities, which affect the growth of the head and face. Dysregulation of cranial neural crest cell (NCC) migration, proliferation, differentiation, and/or cell fate specification have been reported to contribute to CFA. Understanding of the mechanisms through which cranial NCCs contribute for craniofacial development may lead to identifying meaningful clinical targets for the prevention and treatment of CFA. Isolation and culture of cranial NCCs in vitro facilitates screening and analyses of molecular cellular mechanisms of cranial NCCs implicated in craniofacial development. Here, we present a method for the isolation and culture of cranial NCCs harvested from the first branchial arch at early embryonic stages. Morphology of isolated cranial NCCs was similar to O9-1 cells, a cell line for neural crest stem cells. Moreover, cranial NCCs isolated from a transgenic mouse line with enhanced bone morphogenetic protein (BMP) signaling in NCCs showed an increase in their chondrogenic differentiation capacity, suggesting maintenance of their in vivo differentiation potentials observed in vitro. Taken together, our established method is useful to visualize cellular behaviors of cranial NCCs.
Keywords: NCCs (NCCs)Background
Cranial neural crest cells (NCCs) form the face and the anterior part of the head (Jiang et al., 2002; Chai and Maxson, 2006; Mishina and Snider, 2014). Cranial NCCs are one of the subpopulations of NCCs that show unique characteristicis by differentiating into bones and cartilage, while other populations of NCCs do not have this differentiation potential (Stemple and Anderson, 1992; Jiang et al., 2000; Ishii et al., 2012). Cranial NCCs delaminate from the neural tube and migrate into branchial arches (BAs) (Santagati and Rijli, 2003; Kaucka et al., 2016; Yang et al., 2021). Subsequently, they give rise to neural tissues, including peripheral nerve systems but also connective tissues, such as bones, cartilage, and adipose tissues (Santagati and Rijli, 2003). Several signaling pathways are known to regulate differentiation, proliferation, migration, and cell specification for cranial NCCs, which have been reported to develop craniofacial anomalies (CFA), such as cleft palate and craniosynostosis (Teng et al., 2008; Komatsu et al., 2013; He and Soriano, 2013). CFA are a diverse group of pathologic conditions caused by environmental and genetic reasons. Therefore, a deeper understanding of the molecular and cellular mechanisms underpinning cranial NCCs function may result in improved identification, prevention, and treatment of CFA in human patients.
In the 1980s, a method to generate chick-quail chimeras was developed, and cell lineage tracing experiments of neural crest cells during embryonic development became popular (Couly and Dourin, 1985). After establishment of Cre-loxP technologies, transgenic mouse lines expressing Cre recombinase driven by promoters of NCC marker genes, such as Wnt1 and Protein zero (P0) (Wnt1-Cre and P0-Cre lines), facilitated cell lineage tracing during neural crest migration and differentiation (Danielian et al., 1998; Yamauchi et al., 1999, Chen et al., 2017). Using the Cre-loxP technologies, genes of interest are specifically mutated in NCCs to introduce either gain-of-function (GOF) or loss-of-function (LOF) mutations, which significantly deepen our understanding of the mechanisms of craniofacial and trunk development mediated by NCCs. To further gain insight into mechanisms which control proliferation, cell fate specification, and differentiation of cranial NCCs, it is important to establish an in vitro culture system to maintain the character of cranial NCCs.
Bone morphogenetic proteins (BMPs) are a group of growth factors that bind to receptors on the plasma membrane that transduce a signal mediated by SMAD proteins. The levels of BMP-SMAD signaling are important to determine a range of cell behaviors including fate specification; therefore, developing an understanding of the fine-tuning mechanisms of BMP signaling in cranial NCCs is critical. We and others have reported that mutant mice with GOF or LOF of BMP type1A receptor (Bmpr1a) in NCCs display deformities in the craniofacial region (Komatsu et al., 2013; Li et al., 2013; Gu et al., 2014). In addition to that, we have recently reported that when ACVR1 (caAcvr1), another BMP type 1 receptor, is constitutively activated in NCCs of transgenic mice (caAcvr1; P0-Cre mice, hereafter), these formed ectopic cartilage in their face (Yang et al., 2021). Thus, we have proposed that enhanced BMP signaling pushes cranial NCCs to the chondrogenic fate at early embryonic stages.
We established the isolation and culture method for cranial NCCs from the first branchial arch (BA1) at an early embryonic stage, day 10.5 embryos (Figures 1, 2). Isolated cranial NCCs from caAcvr1; P0-Cre mice showed greater chondrogenic differentiation capacity than cranial NCCs from control mice (Figure 3), suggesting that the isolated cranial NCCs from BA1 maintain the differentiation potential observed in vivo. This isolation method is an important tool to analyze molecular and cellular mechanisms of craniofacial development.
Materials and Reagents
1.5 mL tube
Wide bore filtered 1,000 µL pipette tips (Thermo Fisher Scientific, catalog number: 2079G)
35 mm tissue culture-treated dish (SPL Life Sciences, catalog number: 20060)
Sterile Cell Strainer, 40 µm Nylon Mesh (Thermo Fisher Scientific, catalog number: 22363547)
Phosphate buffered saline (PBS), pH 7.4 (Thermo Fisher Scientific, catalog number: 10010023)
Cell-dissociation enzyme, TrypLE Express (Thermo Fisher Scientific, catalog number: 12605028)
Trypan Blue Solution, 0.4% (Thermo Fisher Scientific, catalog number: 15250061)
Advanced DMEM (Thermo Fisher Scientific, catalog number: 12491015)
Ham’s F-12 Nutrient Mix (Thermo Fisher Scientific, catalog number: 11765054)
Penicillin-Streptomycin, 100× (Thermo Fisher Scientific, catalog number: 15140122)
Fetal bovine serum (FBS)-US source, heat inactivated (Denville Scientific, catalog number: FB5001-H)
Fibronectin, human protein, plasma (Thermo Fisher Scientific, catalog number: 33016015)
Bouin’s fixative (Polysciences, catalog number: 16045)
Ammonium hydroxide solution, 28–30% (Sigma-Aldrich, catalog number: 221228)
Alcian blue, 8GX (Sigma-Aldrich, catalog number: A3157)
Acetic acid, glacial (Sigma-Aldrich, catalog number: 695092-2.5L)
Guanidine hydrochloride (Sigma-Aldrich, catalog number: G3272)
Fibronectin stock solution (1 mg/mL) (see Recipes)
100 µg/mL fibronectin in 10% FBS/Advanced DMEM (see Recipes)
10 µg/mL fibronectin in 10% FBS/Advanced DMEM (see Recipes)
4% FBS in Advanced DMEM/Ham’s F-12 (see Recipes)
Solution A for alcian blue staining (see Recipes)
Solution B for alcian blue staining (see Recipes)
1 mg/mL Alcian blue solution (see Recipes)
6M guanidine hydrochloride (see Recipes)
Equipment
Dumont #5 tweezer, tip size 0.10 × 0.06 mm (Roboz, catalog number: RS-5045), sterilized by autoclave
Dumont #7 tweezer, tip size 0.17 × 0.10 mm (Roboz, catalog number: RS-5047), sterilized by autoclave
Micro dissecting scissors, 3.5” straight (Roboz, catalog number: RS-5910), sterilized by autoclave
Stereo microscope, any brand (e.g., Leica M165-FC)
Centrifuge (Thermo Sorval Legend Micro 21)
Class II A/B3 Biological safety cabinet
CO2 incubator, NAPCO SERIES 8000DH
Varioskan Flash Spectral Scanning Multimode Reader (Thermo Scientific)
Procedure
文章信息
版权信息
© 2022 The Authors; exclusive licensee Bio-protocol LLC.
如何引用
Readers should cite both the Bio-protocol article and the original research article where this protocol was used:
- Ueharu, H., Yang, J., Komatsu, Y. and Mishina, Y. (2022). Isolation and Culture of Cranial Neural Crest Cells from the First Branchial Arch of Mice. Bio-protocol 12(7): e4371. DOI: 10.21769/BioProtoc.4371.
- Yang, J., Kitami, M., Pan, H., Toda-Nakamura, M., Zhang, H., Liu, F., Zhu, L., Komatsu, Y. and Mishina, Y. (2021). Augmented BMP signaling commits cranial neural crest cells to a chondrogenic fate by suppressing autophagic b-catenin degradation. Sci Signal 14(665): eaaz9368.
分类
发育生物学 > 细胞生长和命运决定 > 分化
干细胞 > 成体干细胞 > 神经干细胞
细胞生物学 > 细胞分离和培养 > 细胞分化
您对这篇实验方法有问题吗?
在此处发布您的问题,我们将邀请本文作者来回答。同时,我们会将您的问题发布到Bio-protocol Exchange,以便寻求社区成员的帮助。