

A Method to Induce Brown/Beige Adipocyte Differentiation from Murine Preadipocytes
一种从鼠前脂肪细胞诱导棕色/米色脂肪细胞分化的方法
发布: 2021年12月20日第11卷第24期 DOI: 10.21769/BioProtoc.4265 浏览次数: 5029
评审: Ralph Thomas BoettcherClara Lubeseder-MartellatoKossay ZaouiAnonymous reviewer(s)

参见作者原研究论文
The authors used this protocol in:

Dec 2020
Advertisement










Abstract
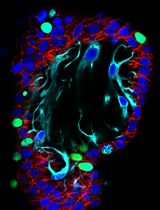
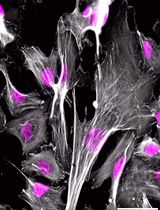
Adipocytes exhibit different morphological and functional characteristics, depending on their anatomical location, developmental origin, and stimulus. While white adipocytes tend to accumulate energy as triglycerides, brown and beige adipocytes tend to direct carbon sources to fuel thermogenesis. White and beige adipocytes originate from common progenitor cells, which are distinct from brown adipocyte precursors. Having a method to study white vs. beige vs. brown adipocyte differentiation may help to unveil the mechanisms driving distinct adipogenic programs. Preadipocytes can be cultured and differentiated in vitro using a combination of compounds to stimulate adipogenesis. Here, we describe and compare protocols designed to stimulate adipocyte differentiation and induce brown/beige-like or white-like characteristics in differentiating adipocytes. The protocols consist in exposing murine preadipocytes to pharmacological stimuli aimed at triggering adipogenesis and inducing (or not) a thermogenic gene expression program. After 8 days of differentiation with a pro-browning cocktail, immortalized preadipocytes isolated from interscapular brown fat (9B cells) or inguinal white fat (9W cells) from the same mouse expressed higher levels of brown/beige adipocyte markers (e.g., Ucp1) and pan-adipocyte differentiation markers (e.g., Pparg, Cebpa and aP2) when compared to the same cells differentiated with a cocktail that lacked brown/beige adipogenic inducers (i.e., rosiglitazone, T3, and indomethacin). Consistent with a higher thermogenic potential of brown vs. beige adipocytes, differentiated 9B cells expressed higher Ucp1 levels than differentiated 9W cells. This simple protocol may help researchers to understand mechanisms of adipogenesis and how adipocytes become thermogenic.
Background
Obesity is characterized by excess body fat and represents a risk factor for several chronic non-transmissible diseases (World Health Organization, 2015). The activation of brown adipose tissue (BAT) and the recruitment of newly formed beige adipocytes in white adipose tissue (WAT) leads to increased thermogenesis and higher energy expenditure, thus representing a promising intervention to treat obesity and its complications. Brown adipocytes contain multiple lipid droplets in their cytoplasm. They are also filled with mitochondria that promote energy dissipation in the form of heat, by leaking protons across the inner mitochondrial membrane, thus uncoupling oxidative phosphorylation from ATP synthesis. This mechanism is mediated by uncoupling protein 1 (UCP1), in a process called non-shivering thermogenesis (Nedergaard et al., 2001; Cannon and Nedergaard, 2004). White adipocytes have a single and large lipid droplet, few mitochondria, and produce many types of adipokines (Rosen and Spiegelman, 2014). Adipocytes with increased thermogenic capacity also appear in WAT depots, under states such as cold exposure, exercise, and caloric restriction (Cinti et al., 2005; Wu et al., 2012; Fabbiano et al., 2016), and are called beige adipocytes. Although beige adipocytes are functionally similar to brown adipocytes, these two types of thermogenic cells differ in their developmental origin (Ikeda et al., 2018). In fact, beige adipocytes share common progenitor cells with white adipocytes (Ikeda et al., 2018). The process of beige adipogenesis is often called browning, and normally occurs in subcutaneous fat depots. Together, these observations suggest that cell extrinsic and intrinsic factors contribute to determine the fate of adipocyte differentiation. Understanding these factors may help elucidating adipogenesis and lead to the identification of new therapeutic targets (Vegiopoulos et al., 2017). Here, we used murine preadipocyte cell lines isolated from different fat depots to characterize and compare protocols to promote brown, beige, or white adipocyte differentiation. These protocols were based on previous studies (Morrison and Farmer, 1999; Fasshauer et al., 2001; Tseng et al., 2004), although they have been slightly modified in our laboratory, where they have been replicated and optimized to apply basic procedures, budget-friendly reagents, and minimal infrastructure.
Materials and Reagents
12-well Cell Culture Plate (Corning, 12-565-321)
0.22 μm sterile filter
9B preadipocytes: these cells were isolated from the interscapular brown fat of a Dicerlox/lox; aP2-Cre-ERT2 mouse and immortalized using SV-40-large T antigen as described before (Mori et al., 2012 and 2014). They behave like wild type cells unless 4-hydroxytamoxifen is added to the medium
9W preadipocytes: these cells were isolated from the subcutaneous white fat of the same mouse used to isolate 9B cells. These cells were immortalized using SV-40-large T antigen and also behave like wild type cells unless 4-hydroxytamoxifen is added to the medium
20 nM insulin* (Sigma-Aldrich, catalog number: I6634, powder storage temperature -20°C. Store at 4°C after dilution)
1 nM triiodothyronine (T3)* (Sigma-Aldrich, catalog number: T2877, storage temperature -20°C)
0.5 mM 3-Isobutyl-1-methylxanthine (IBMX)* (Sigma-Aldrich, catalog number: I5879, storage temperature -20°C)
1 μM dexamethasone* (Sigma-Aldrich, catalog number: D4902, storage temperature -20°C)
0.125 mM indomethacin* (Sigma-Aldrich, catalog number: I7378, storage temperature -20°C)
2.8 μM rosiglitazone* (Sigma-Aldrich, catalog number: R2408, storage temperature -20°C)
Fetal bovine serum (FBS) (Thermo Fisher Scientific, catalog number: 12657-029, storage temperature -20°C)
Penicillin-Streptomycin (Pen/Strep) (Thermo Fisher Scientific, catalog number: 15140122, storage temperature -20°C)
Dulbecco’s modified Eagle’s medium (DMEM), high glucose (Life Technologies, InvitrogenTM, catalog number: 11965, storage temperature 4°C)
1 mM insulin stock solution (see Recipes)
10 μM T3 stock solution (see Recipes)
0.25 M IBMX stock solution (see Recipes)
1 mM dexamethasone stock solution (see Recipes)
0.125 M indomethacin stock solution (see Recipes)
2.8 mM rosiglitazone stock solution (see Recipes)
Maintenance medium (see Recipes)
Differentiation medium day 2 (see Recipes)
Differentiation medium day 4 (see Recipes)
Differentiation medium day 6 (see Recipes)
*Final concentration
Equipment
Heating block
Tissue culture incubator (37°C, 5% CO2, Sheldon Manufacturing, Inc.)
Water bath (Fisher Scientific, FS-205)
Software
Graphpad Prism 9® software
Procedure
文章信息
版权信息
© 2021 The Authors; exclusive licensee Bio-protocol LLC.
如何引用
Readers should cite both the Bio-protocol article and the original research article where this protocol was used:
- Rocha, A. L., Guerra, B. A., Boucher, J. and Mori, M. A. (2021). A Method to Induce Brown/Beige Adipocyte Differentiation from Murine Preadipocytes. Bio-protocol 11(24): e4265. DOI: 10.21769/BioProtoc.4265.
- Rocha, A. L., de Lima, T. I., de Souza, G. P., Correa, R. O., Ferrucci, D. L., Rodrigues, B., Lopes-Ramos, C., Nilsson, D., Knittel, T. L., Castro, P. R., et al. (2020). Enoxacin induces oxidative metabolism and mitigates obesity by regulating adipose tissue miRNA expression. Sci Adv 6(49): eabc6250.
分类
发育生物学 > 细胞生长和命运决定 > 分化
细胞生物学 > 细胞分离和培养 > 细胞分化
生物科学 > 生物技术
您对这篇实验方法有问题吗?
在此处发布您的问题,我们将邀请本文作者来回答。同时,我们会将您的问题发布到Bio-protocol Exchange,以便寻求社区成员的帮助。