

Visualization and Quantitation of Wg Trafficking in the Drosophila Wing Imaginal Epithelium
果蝇翅成虫上皮中Wg运输的可视化和定量研究
发布: 2021年06月05日第11卷第11期 DOI: 10.21769/BioProtoc.4040 浏览次数: 4604
评审: Pradeep Kumar BhaskarLeonardo Gaston GuilgurAnonymous reviewer(s)
参见作者原研究论文
The authors used this protocol in:
Aug 2020
Advertisement










Abstract
Secretory Wnt trafficking can be studied in the polarized epithelial monolayer of Drosophila wing imaginal discs (WID). In this tissue, Wg (Drosophila Wnt-I) is presented on the apical surface of its source cells before being internalized into the endosomal pathway. Long-range Wg secretion and spread depend on secondary secretion from endosomal compartments, but the exact post-endocytic fate of Wg is poorly understood. Here, we summarize and present three protocols for the immunofluorescence-based visualization and quantitation of different pools of intracellular and extracellular Wg in WID: (1) steady-state extracellular Wg; (2) dynamic Wg trafficking inside endosomal compartments; and (3) dynamic Wg release to the cell surface. Using a genetic driver system for gene manipulation specifically at the posterior part of the WID (EnGal4) provides a robust internal control that allows for direct comparison of signal intensities of control and manipulated compartments of the same WID. Therefore, it also circumvents the high degree of staining variability usually associated with whole-tissue samples. In combination with the genetic manipulation of Wg pathway components that is easily feasible in Drosophila, these methods provide a tool-set for the dissection of secretory Wg trafficking and can help us to understand how Wnt proteins travel along endosomal compartments for short- and long-range signal secretion.
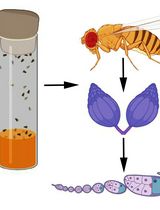
Graphic abstract:

Figure 1. Visualization of extracellular and intracellular Wg trafficking in Drosophila wing imaginal discs. While staining of extracellular Wg without permeabilization exclusively visualizes Wg bound to the extracellular surface (left), Wg uptake and endosomal trafficking can be visualized using an antibody uptake assay (middle). Dynamic Wg release can be visualized by performing a non-permeabilizing staining at a permissive temperature that sustains secretory Wg transport (right).
Background
The developing epithelium of the Drosophila wing imaginal disc (WID) is a well-established model system to study the intracellular trafficking of Wnt proteins. Imaginal discs are epithelial sac-like structures of Drosophila larvae that give rise to adult body parts during pupal transformation. Development of the fly wing from WID depends on the correct formation of several wing axes. To this end, several morphogen gradients are established. The dorsal-ventral wing axis is defined by Wg (Drosophila Wnt-1) secretion from a narrow cell stripe of Wg-expressing cells along the dorsal-ventral boundary. Upon secretion, Wg travels across the WID epithelium over short and long distances to activate target gene transcription in a dose-dependent manner. To ensure Wg gradient formation and proper wing development, Wg secretion and spread from the polarized WID epithelial monolayer are tightly regulated.
Upon translation, Wg is lipid-modified in the endoplasmic reticulum (ER) and transported via the Golgi toward the apical plasma membrane with its cargo receptor Evi/Wls (Bänziger et al., 2006; Bartscherer et al., 2006). While Wg can be transferred to neighboring cells by cell-cell contacts (Alexandre et al., 2014), long-range Wg secretion and signal transduction depend on Wg entry into the endosomal system (Strigini and Cohen, 2000; Pfeiffer et al., 2002). To this end, Wg re-enters the cell after apical presentation through a dynamin-independent endocytic route that delivers Wg into endosomal compartments (Hemalatha et al., 2016). Basal routes for secondary Wg secretion have also been described (Yamazaki et al., 2016) and Wg colocalizes with Rab4-recycling endosomes on the apical WID cell side (Gao et al., 2017; Linnemannstöns et al., 2020; Witte et al., 2020). Apical, secondary Wg release from Rab4-recycling endosomes could therefore fuel the Wg gradient independently of apicobasal transport (Witte et al., 2020). Consequently, interference with the apical release of Wg significantly hinders Wg gradient formation (Chaudhary and Boutros, 2019; Linnemannstöns et al., 2020).
The visualization of Wg trafficking to the cell surface and in post-endocytic endosomal compartments is an important tool to understand how the long- and short-range Wg signal is generated. Here, we present three main methods that have been established to unravel which of the proposed secretory routes feed the Wg signal: (1) the analysis of steady-state extracellular Wg levels [initially established by Strigini and Cohen (2000)]; (2) the visualization of intracellular Wg transport after endocytic uptake [modified from Hemalatha et al. (2016)]; and (3) the visualization of Wg recycling (Witte et al., 2020) (Figure 1). All three methods are based on immunofluorescence staining of dissected Drosophila WID, but different approaches are used to visualize Wg in the respective WID compartment (Figure 2). Briefly, while (1) and (3) make use of non-permeabilizing staining protocols, albeit at different temperatures, to specifically visualize surface bound extracellular Wg, (2) uses the endocytic uptake of antibody-labeled Wg to visualize its endosomal transport. Using UAS/Gal4-driven gene expression in combination with an engrailed promotor to drive genetic manipulations specifically in posterior WID allows the direct quantitation of anterior and posterior phenotypes within the same WID.
Multiple intracellular trafficking routes have been proposed to contribute to Wg signal transfer. It is therefore especially interesting to see how extracellular and intracellular Wg distributions react to interference with different aspects of the Wg trafficking machinery. As genetic manipulations are comparatively easy in Drosophila, the WID provides an appropriate means to probe this system. We hope that the methods presented herein contribute to gaining further insight into the evolutionarily conserved secretory Wg/Wnt trafficking pathway in order to fully understand how Wnt signals can be transferred over short and long distances. Moreover, these protocols are not limited to Wg and can be used to investigate the trafficking of other morphogens, such as Hedgehog (Hh) or Decapentaplegic (Dpp).

Figure 2. Overview of the different staining approaches. After WID dissection, extracellular Wg staining is performed on ice without permeabilization. The Wg uptake assay is performed at 22°C and includes acid washing to remove extracellularly bound signal, as well as permeabilization. Wg recycling is monitored at 22°C in the absence of permeabilization. After staining, WID samples are again dissected and mounted for downstream imaging and quantitation.
Materials and Reagents
Drosophila vials (Kisker Biotech GmbH, catalog number: 789013)
Plugs (Flugs wide plastic vials, Kisker Biotech GmbH, catalog number: 789035)
1.5 ml Reaction tubes (Eppendorf 3810X, catalog number: 0030 125.150)
Microscope slides (LABSOLUTE, catalog number: 7 695 002)
Cover slips (Roth, catalog number: H878)
Ice and ice bucket
Drosophila third instar larvae
Fetal bovine serum (FBS) (Sigma-Aldrich, catalog number: S0615)
Schneider’s insect medium (Sigma-Aldrich, catalog number: S0146)
Shields and Sang M3 insect medium (Sigma-Aldrich, catalog number: S3652)
16% Paraformaldehyde (Electron Microscopy Sciences, catalog number: 15710-S)
Mouse anti-Wg (Developmental Studies Hybridoma Bank, catalog number: 4D4)
Goat anti-mouse Alexa Fluor 647 (1:500, Invitrogen, catalog number: A21236)
Hoechst 33342 (1:500, Thermo Scientific, catalog number: 62249)
Mowiol (Calbiochem, catalog number: 475904)
Corn meal (Spielberger Mühle, catalog number: SP061388)
Soy flour (Spielberger Mühle, catalog number: SP061388)
Dry yeast (Bierhefe Pulver, Cenovis)
Malt extract (Lindenmeyer, Demeter)
Sugar beet syrup (Bauckhof, Demeter)
Agar-Agar (Roth, catalog number: 5210.2)
Propionic acid (Roth, catalog number: 6026.3)
Nipagin (700 ml 99% ethanol) (Roth, catalog number: 9065.4)
Nipagin (C8H8O3) (Fluka, catalog number: 54752-1KG-F)
NaCl (Roth, catalog number: 3957.2)
KCl (Roth, catalog number: 6781.1)
Na2HPO4·2H2O (Roth, catalog number: 4984.1)
KH2PO4 (Fluka, catalog number: 15645940)
Triton X-100 (Roth, catalog number: 3051.4)
Glycine (Roth, catalog number: 3908.3)
HCl (Roth, catalog number: K025.1)
Fly food (see Recipes)
PBS (see Recipes)
PBTx (see Recipes)
Glycine- HCl buffer (see Recipes)
Mowiol (see Recipes)
Equipment
Drosophila incubator (RUMED, type 3201)
Brush (e.g., davinci, size 2)
Forceps (Roth, catalog number: K342.1)
Fluorescence stereomicroscope (depending on genetic set-up) (e.g., SZX12, equipped with U-RFL-T, Olympus)
Dissection microscope such as Stemi 2000 (Zeiss) equipped with light source (e.g., KL-200, Zeiss)
Confocal microscope such as LSM780 confocal laser-scanning microscope (Zeiss), equipped with Plan Neofluar 63×/oil NA 1.4 and PlanNeofluar 10× NA 0.3 objectives (Zeiss)
Thermoblock (Eppendorf, Thermomixer compact)
Rocker (Heidolph, POLYMAX 1040)
Software
Zen Black (Zeiss), Version 14.0.0.0 (or alternative software for image aquisition)
Fiji/ImageJ (NIH) (Schneider et al., 2012; Rueden et al., 2017), Version 1.53c (for image quantitation)
Microsoft Excel, Microsoft Office Professional Plus 2016 (or alternative software for data analysis)
Procedure
文章信息
版权信息
© 2021 The Authors; exclusive licensee Bio-protocol LLC.
如何引用
Witte, L., Linnemannstöns, K., Honemann-Capito, M. and Gross, J. C. (2021). Visualization and Quantitation of Wg Trafficking in the Drosophila Wing Imaginal Epithelium. Bio-protocol 11(11): e4040. DOI: 10.21769/BioProtoc.4040.
分类
细胞生物学 > 细胞成像 > 共聚焦显微镜
发育生物学 > 细胞信号传导 > 配体
发育生物学 > 形态建成 > 细胞结构
您对这篇实验方法有问题吗?
在此处发布您的问题,我们将邀请本文作者来回答。同时,我们会将您的问题发布到Bio-protocol Exchange,以便寻求社区成员的帮助。