

Multitarget Immunohistochemistry for Confocal and Super-resolution Imaging of Plant Cell Wall Polysaccharides
植物细胞壁多糖共聚焦和超分辨成像的多靶点免疫组织化学研究
(*contributed equally to this work) 发布: 2020年10月05日第10卷第19期 DOI: 10.21769/BioProtoc.3783 浏览次数: 7980
评审: Ayelign M. AdalSara Posé Anonymous reviewer(s)
参见作者原研究论文
The authors used this protocol in:

Feb 2020
Advertisement











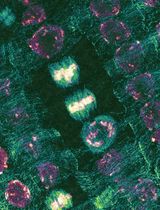
Abstract
The plant cell wall (PCW) is a pecto-cellulosic extracellular matrix that envelopes the plant cell. By integrating extra-and intra-cellular cues, PCW mediates a plethora of essential physiological functions. Notably, it permits controlled and oriented tissue growth by tuning its local mechano-chemical properties. To refine our knowledge of these essential properties of PCW, we need an appropriate tool for the accurate observation of the native (in muro) structure of the cell wall components. The label-free techniques, such as AFM, EM, FTIR, and Raman microscopy, are used; however, they either do not have the chemical or spatial resolution. Immunolabeling with electron microscopy allows observation of the cell wall nanostructure, however, it is mostly limited to single and, less frequently, multiple labeling. Immunohistochemistry (IHC) is a versatile tool to analyze the distribution and localization of multiple biomolecules in the tissue. The subcellular resolution of chemical changes in the cell wall component can be observed with standard diffraction-limited optical microscopy. Furthermore, novel chemical imaging tools such as multicolor 3D dSTORM (Three-dimensional, direct Stochastic Optical Reconstruction Microscopy) nanoscopy makes it possible to resolve the native structure of the cell wall polymers with nanometer precision and in three dimensions.
Here we present a protocol for preparing multi-target immunostaining of the PCW components taking as example Arabidopsis thaliana, Star fruit (Averrhoa carambola), and Maize thin tissue sections. This protocol is compatible with the standard confocal microscope, dSTORM nanoscope, and can also be implemented for other optical nanoscopy such as STED (Stimulated Emission Depletion Microscopy). The protocol can be adapted for any other subcellular compartments, plasma membrane, cytoplasmic, and intracellular organelles.
Background
PCW is an intricate material, which is durable but also undergoes constant structural changes in response to internal and external stimuli such as tissue expansion or pathogen attack. Nevertheless, how these seemingly contradictory features, mechanical strength and structural adaptation, cooperate, remains an unresolved question in plant cell biology. PCW contains cellulose, hemicellulose, different variants of pectin and various proteins, the architecture of which is highly organized (Peaucelle, 2018). The pectin family is composed of several polymers. The most abundant, homogalacturonan, can be demethylated after cell wall insertion. This change in chemistry is a significant step in the process of cell elongation, differentiation, and directional growth (Peaucelle et al., 2012). Several lines of evidence suggest that morphogenesis and cell differentiation are dependent on local changes in cell wall chemistry and cell wall polymer organization (Yang et al., 2016; Anane et al., 2017; Zhao et al., 2019; Haas et al., 2020). Therefore, detailed knowledge of PCW components architecture is essential for understanding plant growth. Historically, the cell wall structure has been studied using biochemical methods that involve disintegrating the tissue and destroying the native organization of its polymers (Höfte and Voxeur, 2017). Other imaging modalities, such as electron microscopy (EM) (Anane et al., 2017) and atomic force microscopy (AFM) (Zhang et al., 2017) are used for in situ cell wall observation, but these techniques often lack chemical contrast, providing only correlative quantification. Multicolor immunohistochemistry (IHC) can reveal multiple targets within a tissue section, and their spatial organization resolved in three dimensions. Despite its widespread use in the cell biology field, multicolor IHC is not yet a standard tool for studies of the cell wall. A broad palette of antibodies and probes against cell wall targets, coupled with multicolor IHC, together with the high resolving power of dSTORM (< 40 nm), permits quantitative in situ chemical analysis of cell wall nanostructure. Other imaging techniques for cell wall analysis on tissue cuts exist, such as Raman (Wightman et al., 2019) and FTIR (Mravec et al., 2017; Cuello et al., 2020). These techniques are based on the characteristic absorption/transmission of different chemical components and can measure some changes in cell wall composition at the cellular level but can lack sensitivity, and their ability to observe changes at the subcellular level is severely limited.
The dSTORM permits localization of biomolecules with the precision of 5-10 nm; however, the final dSTORM resolution, typically around 40 nm, is limited by the size of the antibody complex (~15-30 nm). The immunogold Electron Microscope (iEM) is also used in combination with ICH to study cellular structures at high resolutions. IEM is comparable to dSTORM resolution and is also limited by the antibody complex size. The size of the nanogold particle defines iEM contrast and resolution; > 1 nm gold nanoparticles are available; however, such a small particle limits the contrast, and larger particles are often used. IEM, contrasted with dSTORM, has several additional drawbacks: (1) the primary antibodies probed with protein-A (or G) gold complexes do not penetrate through the resin-embedded sample, and only recognize the surface epitopes, although the serial and ultra-thin cryo-sectioning technique can resolve this problem (50 nm, -120 °C [Slot, 1989]); (2) samples are mostly single-labeled; yet by using different nanoparticle size, two or three epitopes can be tagged, but it requires the technical experiences of ultra-thin cryo-sectioning technique (Slot and Geuze, 2007); (3) IEM has low labeling and detection efficiency (3-5 orders of magnitude less than dSTORM (Majda et al., 2017; Haas et al., 2020), which is also related to the fact, that only the surface epitopes are labeled. Multicolor 3D dSTORM nanoscopy can, therefore, provide unprecedented insights into the nanoarchitecture of the native-structure of the cell wall polysaccharides, beyond the applicability of the aforementioned techniques. DSTORM permits a quantitative three-dimensional (3D) nanoimaging of cells and tissues (Heilemann et al., 2008; Huang et al., 2008; Van De Linde et al., 2011; Sydor et al., 2015; Xu et al., 2018). Applied mainly to cellular systems, it unveiled new structural organizations of proteins, e.g., synaptic nanodomains, trans-synaptic nanocolumns, DNA recombinase nanofilaments, nuclear envelope pore structure, cytoskeleton, mitochondria, adhesion complexes, and chromatin transcriptional landscape (Shroff et al., 2008; Shim et al., 2012; Löschberger et al., 2012; Jakobs and Wurm, 2014; Prakash et al., 2015; Boettiger et al., 2016; Sellés et al., 2017; Dlasková et al., 2018; Haas et al., 2018a and 2018b; Pan et al., 2018; Xia et al., 2019; Chen et al., 2020; Wäldchen et al., 2020). Yet, due to the limitation of single-cell model systems requiring tissue-level imaging, its utilization in plant science is almost absent (Liesche et al., 2013; Komis et al., 2015; Haas et al., 2020). Here we present a detailed protocol for the sample preparation compatible with a standard confocal microscope and a dSTORM nanoscope on thin plant tissue sections.
Materials and Reagents
- 8-well ibidi µ-slides for dSTORM sample preparation and imaging (Ibidi, catalog number: 80827)
- Biopsy cassette 42 x 28 x 6 mm (Leica Biosystems, ID: IP-Biopsy-cassette-III)
- Metal base mold (Leica Biosystems, ID: metal-base-molds)
- Charged slides by poly-L-lysine treatment for confocal imaging sample preparation and imaging such as ProbeOn Plus (Fisher Scientific catalog number: 22-230-900 or PolysineTM Microscope Adhesion Slides (Thermo Scientific catalog number: J2800AMNZ)
- Aluminum foil (you can get it from a local store)
- Coverglass, rectangular, 24 x 60 mm, #1.5 thickness (Knittel Glass, catalog number: 425-2460)
- Eppendorf Safe-lock Microcentrifuge Tubes, 1.5 ml or 2 ml volume (Eppendorf, catalog numbers: 022-36-320-4 [1.5 ml], 022-36-335-2 [2 ml])
- Cardboard Slide Tray (Heathrow Scientific, catalog number: HD9902)
- Paper towels (Divers/Dutcher, catalog number: 475040)
- Empty pipette tips box for homemade humidified chamber for ibidi µ-slides, such as Adamas-Beta 1,000 µl volume tips
- Nitrile gloves, such as FisherbrandTM Comfort Nitrile Gloves (Fisher Scientific, catalog number: 15642367)
- National DiagnosticsTM HistoclearTM Tissue clearing agent (Fisher Scientific, HS-200-1GAL, CAS number: 5989-27-5)
- Embedding medium paraffin (Leica Biosystems, ID: em-400-embedding-medium-paraffin)
- Oxoid Skim Milk Powder (Thermo Fisher Scientific, catalog number: LP0031B)
- CMB3a Crystalline cellulose-binding module, his-tagged and recombinant CBM protein (PlantProbes, catalog number: CMB3a) (Blake et al., 2006; Hernandez-Gomez et al., 2015), store at -20 °C
- Mouse monoclonal antibody against low esterified homogalacturonan (degrees of methyl-esterification (DM) up to 40%) 2F4 (PlantProbes, catalog number: 2F4) (Liners, Thibault and Van Cutsem, 1992), store at 4 °C
- Rat IgM monoclonal antibody against high-esterified homogalacturonan LM20 (PlantProbes, catalog number: LM20) (Verhertbruggen et al., 2009), store at 4 °C
- Rat IgA monoclonal antibody against partially methyl-esterified epitopes of homogalacturonan (PlantProbes, catalog number: JIM7) (Knox et al., 1990), store at 4 °C
- Rat IgG2a monoclonal antibody against xyloglucan binding preferentially to the XLLG motif of xyloglucan LM24 (PlantProbes, catalog number: LM24) (Tanackovic et al., 2016), store at 4 °C
- Anti-His tag polyclonal antibody produced in chicken, Anti-6X-His tag (Abcam, catalog number: ab9107) store at -20 °C
- Anti-His tag polyclonal antibody produced in rabbit (Merck, catalog number: SAB4301134), store at -20 °C
- Rabbit PDM antibody against Mannan was a kind gift by Paul Dupree (Handford et al., 2003; Yang et al., 2016), store at 4 °C. For availability, please contact raymond.wightman@slcu.cam.ac.uk
- Recombinant and His-tagged CBM4 of Cellulomonas fimi (ATCC 484) endoglucanase C (CBD4N1) produced from E. coli. CBM4 was a kind gift by Harry Gilbert (Johnson et al., 1996; Blake et al., 2006), store at 4 °C. For availability, please contact raymond.wightman@slcu.cam.ac.uk
- Goat anti-rat CF568 secondary antibody (Sigma, catalog number: SAB4600086), store at -20 °C
- Goat anti-mouse CF568 conjugated F(ab)’2 secondary antibody fragment (Sigma, catalog number: SAB4600400), store at -20 °C
- Goat anti-mouse F(ab')2 secondary antibody fragment conjugated to Alexa Fluor 647 (Stratech Scientific, catalog number: 115-607-003-JIR), store at 4 °C
- Donkey anti-mouse F(ab')2 secondary antibody fragment conjugated to Alexa Fluor 647 (Abcam, catalog number: ab181292), store at 4 °C
- Donkey anti-Chicken F(ab’)2 secondary antibody fragment conjugated to Alexa Fluor 647 (Jackson immunoresearch, catalog number: 703-606-155), store at 4 °C
- Goat anti-mouse F(ab')2 secondary antibody fragment conjugated to ATTO488 (Hypermole, catalog number: 2402-0.5MG), store at -20 °C
- Goat anti-mouse Alexa 488 conjugated secondary antibody (Sigma, catalog number: SAB4600387), store at 4 °C
- Donkey anti-rat F(ab')2 secondary antibody fragment conjugated to Alexa Fluor 647 (Abcam, catalog number: ab150151), store at 4 °C
- ProLong Gold Antifade Mountant (Thermo Ficher Scientific, catalog number: P36934), store at 4 °C
- Ammonium chloride (NH4Cl) (Sigma-Aldrich, catalog number: 254134), store at RT
- Poly-L-lysinesolution 0.1% (w/v) in H2O (Merck, catalog number: P8920), store at RT
- Ethanol absolute anhydrous (CARLO ERBA reagents, catalog number: 4146082), store at RT
- Acetic Acid ≥ 96% (AnalaR NORMAPUR, catalog number: 20099.324), store at RT
- Formaldehyde (Sigma-Aldrich catalog number: F1635), store at RT
- Arabidopsis inflorescence meristem fixed at 1 cm length stage, and cotyledon of 3 days-old seedling, leaf rachis of star fruit (Supplemental Figure 1AG), immature maize leaf blade at 6th node (Supplemental Figure 1AF)
- Nail polish (you can get it from a local drugstore)
- Pectolyase for enzymatic extraction of pectins (Sigma, catalog number: P3026 or Magazyme,catalog number: E-PLYCJ)
- Calcofluor White Stain for general cell wall staining (Sigma Aldrich, catalog number: 18909-100ML-F)
- 2F4 Buffer (T/Ca/S buffer final concentration) (see Recipes), store at RT
- FAA solution (see Recipes), store at RT
- Formaldehyde diluted in 10x 2F4 buffer (see Recipes)
- Citrate-Phosphate Buffer for Pectolyase incubation (pH 4.8) (see Recipes)
Equipment
- Dispensable microtome knife (Microm Microtech, catalog number: F/MM35p)
- Micro tweezer (Ideal-tek, catalog number: 5-SA)
- Glass Rectangular 250 ml Coplin Staining Jar, with Lid (Wheaton, catalog number: 900620)
- FisherbrandTM Microscope Slide Box for homemade humidified chamber for Polysine microscope slides (Fisher Scientific, catalog number: 22363400)
- Brush for the capture of serial cuts obtained by the microtome (Supplemental Figure 1K, any fine brush from local stationery store, such as Etude, P10531.00, #2)
- HistoCore Arcadia Heated Paraffin Embedding Station (Leica, ID:14039357258)
- Leica EG F Electric Heatable Forceps
- Microtome (Leica, model: RM2265)
For more information, see user’s manual. - Fridge 4 °C
- Incubation chamber, such as Selecta (set at 60 °C), airflow not necessary
- Laboratory freezer (-20 °C), such as Kirsch FROSTER LABO 330 ULTIMATE
- Electronic laboratory heating plate (BIO-OPTICA Milano SpA, catalog number: 40-300-300)
- Laboratory chemical fume hood
- Any Fluorescence Microscope equipped with four laser lines for the detection in UV (405 nm), green (488 nm), red (561 nm) and far red (633 nm) (here we use Zeiss LSM 710, Zeiss Oberkochen Germany)
Brochure available here.
Specification- Stands: Inverted (Axio Observer Z1 with side port port)
- Z drive and XY stage (option): Motorized XY-scanning stage, with Mark & Find function (xyz) and Tile Scan (mosaic scan); the smallest increments 1 μm
- Objectives: x10, x25, x40, x63, x100
- Lasers: Argon laser (458, 488, 514 nm), HeNe laser (633 nm), diode laser 405 nm, and DPSS laser 561 nm
- 405 nm for Calcofluor White
- 488 nm for Alexa 488 and ATTO 488
- 561 nm for CF568 and Alexa 568
- 633 nm for Alexa 647
- Scanning Module
Model: Scanning module with 32 spectral detection channels (QUASAR)
Scanners: Two independent, galvanometric scan mirrors with ultra-short line and frame fly back.
Scanning resolution: 4 x 1 to 6,144 x 6,144 pixels
Scanning speed: 8 frames/sec with 512 x 512 pixels.
Number of fluorescence-spectral detectors: 2
Bright field transmission detector: Installed - Software
Standard Software: ZEN2010
Optional Softwar: Image J or Fiji
Computer specification: HP Z800 Workstation, 64-bit Windows 7 Ultimate 2009, 24 Gb RAM, Intel® Xeon® CPU, X5650, Two processors 2.67 GHz, 2.66 GHz
Wild-field microscope (Nikon N-STORM)
Software
- Grafeo (Custom made software for dSTORM data analysis and visualization, https://github.com/inatamara/Grafeo-dSTORM-analysis- (Haas et al., 2018b)
- Fiji (https://imagej.net/Fiji/Downloads)
Procedure
文章信息
版权信息
© 2020 The Authors; exclusive licensee Bio-protocol LLC.
如何引用
Readers should cite both the Bio-protocol article and the original research article where this protocol was used:
- Haas, K. T., Rivière, M., Wightman, R. and Peaucelle, A. (2020). Multitarget Immunohistochemistry for Confocal and Super-resolution Imaging of Plant Cell Wall Polysaccharides. Bio-protocol 10(19): e3783. DOI: 10.21769/BioProtoc.3783.
- Haas, K. T., Wightman, R., Meyerowitz, E. M. and Peaucelle, A. (2020). Pectin homogalacturonan nanofilament expansion drives morphogenesis in plant epidermal cells. Science 367(6481): 1003-1007.
分类
植物科学 > 植物细胞生物学 > 细胞成像
发育生物学 > 形态建成 > 细胞结构
细胞生物学 > 细胞成像 > 固定组织成像
您对这篇实验方法有问题吗?
在此处发布您的问题,我们将邀请本文作者来回答。同时,我们会将您的问题发布到Bio-protocol Exchange,以便寻求社区成员的帮助。