

Measurement of Acid Ecto-phosphatase Activity in Live Leishmania donovani Parasites
杜氏利什曼原虫中酸性胞外磷酸酶活性的活体检测
发布: 2019年10月05日第9卷第19期 DOI: 10.21769/BioProtoc.3384 浏览次数: 5278
评审: Alexandros AlexandratosCristiane França da SilvaRon Saar Dover
参见作者原研究论文
The authors used this protocol in:

May 2015
Advertisement










Abstract
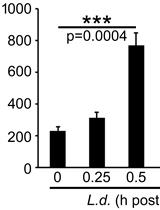
Acid ecto-phosphatases are enzymes that hydrolyze phosphomonoesters in the acidic pH range with their active sites facing the extacellular medium. Their activities can be measured in living cells. In bacteria and protozoan pathogens, acid ecto-phosphatases have been associated with the survival of intracellular pathogens within phagocytes through inhibition of the respiratory burst, suggesting that they act as virulence factors. Extracellular acid phosphatase activity in Leishmania (L.) donovani has been associated with the degree of promastigote virulence/infectivity. The levels of acid ecto-phosphatase activity in different Leishmania sp or even strains of the same species vary and this has been linked to their virulence. It may also be related to their ability to survive and multiply in the insect host.
Acid phosphatase enzymatic activity can be measured in crude membrane fractions and in membrane fractions enriched in plasma membrane, however, in these cases, the intracellular acid phosphatases, mainly localized in lysosomes, contribute to the final result. Therefore, measuring phosphatase activity at the surface of live cells in acidic pH range is the only accurate way to measure acid ecto-phosphatase activity. This assay is performed at 25 °C or 37 °C for 30 min using as substrate the generic phosphatase substrate p-nitrophenyl phosphate (pNPP), in a citrate buffer, with or without sodium tartrate (L(+)-tartaric acid), as histidine acid phosphatases are classified according to their sensitivity to tartate inhibition. The steps of the protocol consist of pelleting cells in suspension, in this case Leishmania promastigotes, washing twice with HEPES buffer, resuspending the cells in the substrate reaction mixture and terminating the reaction by the addition of 0.5 N NaOH. The cells are removed by centrifugation and the absorbance of the reaction product (p-nitrophenolate=pNP) in the supernatant is measured at 405 nm. The enzymatic activity (A405 values) is normalized for the mean number of cells/ml used for each independent experiment.
Background
Trypanosomatid protozoan parasites are a diverse group of primitive parasitic organisms (Kaufer et al., 2017). They are both of medical and veterinary importance as many Trypanosomatid species cause serious or fatal diseases in humans and agricultural animals in tropical and subtropical regions worldwide (Reference 16). Virulence factors of these parasites include molecules expressed on their cell surface as well as those secreted or shed into the extracellular medium (Gottlieb and Dwyer, 1981; Fonseca-de-Souza et al., 2008 and 2009). Trypanosomatidae are exposed to diverse environmental stimuli throughout their life cycles in all biological systems they reside. Central events in recognition of the external and internal signals in these cells, which lead to specific responses for adaptation and survival in all these environments, are protein phosphorylation and dephosphorylation events carried out by a multitude of kinases and phosphatases. The trypanosomatid kinome and phosphatome have been recently deciphered (Brenchley et al., 2007; Sadatomi et al., 2013).
Phosphatase activities modulate different aspects of the infection by members of the trypanosomatid family (Neves et al., 2014). Ecto-phosphatases, the enzymes localized in parasite’s surface membranes, are of special interest as they are candidate molecules for the interaction between the parasite and its host and thereby they could be intimately involved in many cellular events implicated in parasite adhesion to the host, protection from the host’s immune response and establishment of infection. Moreover, several alternative functions have been suggested for ecto-phosphatases, such as participation in proliferation, differentiation, adhesion, virulence and infection (Katakura and Kobayashi, 1988; Singla et al., 1992). Little is known about their physiological roles in protozoa parasites (Cosentino-Gomes et al., 2011; Gomes et al., 2011). In particular, acid ecto-phosphatases that hydrolyze phosphomonoesters at acidic pHs, have been associated with the survival of intracellular parasites within phagocytes through inhibition of the respiratory burst (Saha et al., 1985; Remaley et al., 1985; Baca et al., 1993; Reilly et al., 1996; Jungnitz et al., 1998; Saleh and Belisle, 2000; Burtnick et al., 2001; Steenkamp, 2002; Cosentino-Gomes et al., 2009; Aragon et al., 2011; Cosentino-Gomes and Meyer-Fernandes, 2011), suggesting that they may act as virulence factors (Katakura and Kobayashi, 1988; Singla et al., 1992; Papadaki et al., 2015). They may also be related to the ability of parasites to survive and multiply in the insect host (Remaley et al., 1985; Saha et al., 1985; Singla et al., 1992). The field of the acid ecto-phosphatases was reopened with the characterization of the L. donovani acid ecto-phosphatase LdMAcP (Papadaki et al., 2015) and the identification of at least two more open reading frames in the L. donovani genome that could be potentially expressed at the surface membrane of the parasites (Papadaki and Boleti. data not published).
Therefore assaying acid phosphate activity at the surface of live cells is a putative diagnostic parameter for virulence of Leishmania parasite strains as well as for the evaluation of the expression of the ecto-phosphatases under study at the surface of transgenic parasite strains overexpressing them.Acid phosphatase enzymatic activity can be measured in crude membrane fractions and in membrane fractions enriched in plasma membrane (Glew et al., 1982; Shakarian et al., 2002), however, in these cases, the intracellular acid phosphatases, mainly localized in lysosomes, contribute to the final result.
The protocol provided herein was used to measure acid ecto-phosphatase activity in live Leishmania promastigotes but it could be used to detect and quantitate acid ecto-phosphatase activity in all suspension cells of prokaryotic or eukaryotic origin.
Our method was adapted from previously described methodologies (Remaley et al., 1985; Saha et al., 1985; Katakura and Kobayashi, 1988; Singla et al., 1992) for assaying ecto-phosphatases in other organisms.
Materials and Reagents
- Plastic cuvettes for spectrophotometric analysis (i.e., Sigma-Aldrich, catalog number: C5677)
- Plastic flasks for parasite culture 25 and 50 cm2
- Plastic pipette tips (for 20 μl, 200 μl and 1 ml pipettes)
- Sterile plastic pipettes (1 ml, 2 ml, 5 ml and 10 ml, from any company)
- Falcon plastic tubes 15 and 50 ml (from any company)
- Plastic Eppendorf tubes 1.5 ml (from any company)
- Gilson or Eppendorf pipettes (or from any other company) for 1 μl, 20 μl, 200 μl and 1 ml volumes
- L. tarentolae promastigotes (LEXSY host P10, Jena Biosciences, catalog number: LT-101)
- pLEXSY-sat2 vector (Jena Biosciences, catalog number: EGE-274)
- Brain Heart Infusion (Lexsy Broth BHI, Jena Biosciences, catalog number: ML-412S), stored at 4 °C when diluted in sterile dH2O
- Hemin (Jena Biosciences, catalog number: ML-108L), stored at RT
- L-biopterin (Cayman, catalog number: 10007662), stored at -20 °C
- Nourseothricin (LEXSY NTC, Jena Bioscience, catalog number: AB-102L). Prepare solution of 10 mg/ml with sterile dH2O, store in aliquots of 100 μl at -20 °C and after thawing at 4 °C
- RPMI 1640-GlutaMax (Thermo Fisher, catalog number: 61870-010), stored at 4 °C
- Schneider’s (Sigma-Aldrich, catalog number: S-9895), stored at 4 °C
- hiFBS: FBS (From Gibco, Thermo Fisher, Biosera or any other company tested), stored at -20 °C, inactivated at 56 °C for 30 min before added to the culture medium
- Penicillin-Streptomycin 10,000 U/ml (100x concentrated, Thermo Scientific, catalog number: 15140-122 or from any other company tested), stored at -20 °C
- HEPES 1 M (Thermo Fisher Scientific, catalog number: 15630080), stored at 4 °C
- 4-Nitrophenyl phosphate disodium salt hexahydrate, pNPP (Sigma-Aldrich, catalog number: P4744), powder stored at -20 °C. Prepare solution of 10 mM fresh for each experiment
- 90 mM citrate buffer at pH 4.8 (25 °C) (Sigma-Aldrich, catalog number: C2488), stored at 2-8 °C
- NaOH 0.5 M solution (Applichem, catalog number: 181692.1211), stored at RT
- Sodium tartrate (5 mM) solution; L(+)-tartaric acid (Merck, catalog number: 100804, or Sigma-Aldrich, catalog number: T109), stored at RT
- 0.4% (w/v) Trypan Blue solution (Gibco, catalog number: 15250061; or Sigma-Aldrich, catalog number: T8154)
- Formalin 100% (37% w/v formaldehyde solution; MERC: catalog number F8775 or from any other comany)
Equipment
- Hemocytometer (Malassez, Marienfeld, catalog number: 0640610)
- Incubator operating at 25 °C for the maintenance of parasite cultures (for example Shel-lab microbiological or CO2 incubator, https://www.sheldonmanufacturing.com/shel-lab-incubators or from any other company)
- Water bath (i.e., ED13, Julabo)
- Analytical balance (i.e., ΑΕ50, Mettler)
- Spectrophotometer (i.e., SmartSpec Plus Spectrophotometer, 170-2525, Bio-Rad)
- Tabletop centrifuge (i.e., Jouan CR3i multifunction, Thermo)
- Microcentrifuge (i.e., Eppendorf 5417R)
Software
- GraphPad Prism Software 5.01 (GraphPad, San Diego, CA)
Procedure
文章信息
版权信息
© 2019 The Authors; exclusive licensee Bio-protocol LLC.
如何引用
Papadaki, A. and Boleti, H. (2019). Measurement of Acid Ecto-phosphatase Activity in Live Leishmania donovani Parasites. Bio-protocol 9(19): e3384. DOI: 10.21769/BioProtoc.3384.
分类
生物化学 > 蛋白质 > 活性
细胞生物学 > 基于细胞的分析方法 > 酶学测定
您对这篇实验方法有问题吗?
在此处发布您的问题,我们将邀请本文作者来回答。同时,我们会将您的问题发布到Bio-protocol Exchange,以便寻求社区成员的帮助。