- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

A Novel Method of Inducible Directed Evolution to Evolve Complex Phenotypes
(*contributed equally to this work) Published: Vol 12, Iss 20, Oct 20, 2022 DOI: 10.21769/BioProtoc.4535 Views: 3080
Reviewed by: Alessandro DidonnaVasudevan AchuthanChhuttan L MeenaRita Marie Celine Meganck
Original research article
The authors used this protocol in:

Jun 2022
Advertisement


Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics







Abstract
Directed evolution is a powerful technique for identifying beneficial mutations in defined DNA sequences with the goal of improving desired phenotypes. Recent methodological advances have made the evolution of short DNA sequences quick and easy. However, the evolution of DNA sequences >5kb in length, notably gene clusters, is still a challenge for most existing methods. Since many important microbial phenotypes are encoded by multigene pathways, they are usually improved via adaptive laboratory evolution (ALE), which while straightforward to implement can suffer from off-target and hitchhiker mutations that can adversely affect the fitness of the evolved strain. We have therefore developed a new directed evolution method (Inducible Directed Evolution, IDE) that combines the specificity and throughput of recent continuous directed evolution methods with the ease of ALE. Here, we present detailed methods for operating Inducible Directed Evolution (IDE), which enables long (up to 85kb) DNA sequences to be mutated in a high throughput manner via a simple series of incubation steps. In IDE, an intracellular mutagenesis plasmid (MP) tunably mutagenizes the pathway of interest, located on the phagemid (PM). MP contains a mutagenic operon (danQ926, dam, seqA, emrR, ugi, and cda1) that can be expressed via the addition of a chemical inducer. Expression of the mutagenic operon during a cell cycle represses DNA repair mechanisms such as proofreading, translesion synthesis, mismatch repair, and base excision and selection, which leads to a higher mutation rate. Induction of the P1 lytic cycle results in packaging of the mutagenized phagemid, and the pathway-bearing phage particles infect naïve cells, generating a mutant library that can be screened or selected for improved variants. Successive rounds of IDE enable optimization of complex phenotypes encoded by large pathways (as of this writing up to 36 kb), without requiring inefficient transformation steps. Additionally, IDE avoids off-target genomic mutations and enables decoupling of mutagenesis and screening steps, establishing it as a powerful tool for optimizing complex phenotypes in E. coli.
Graphical abstract:

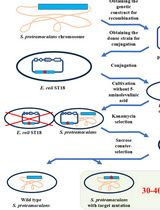
Figure 1. Overview of Inducible Directed Evolution (IDE). Pathways of interest are cloned into a P1 phagemid (PM) backbone and transformed into a strain ofE. coli containing MP (diversification strain). The mutagenesis plasmid is induced to generate mutations. Phage lysate is produced and used to infect a strain that expresses the phenotype of interest (screening/selection strain). The resulting strain library is screened to identify those with improved properties. Narrowed-down libraries can then go through another IDE cycle by infecting a fresh diversification strain.
Background
Complex phenotypes are defined as those that result from the combined action of multiple genes. Frequently, the genes that most limit these phenotypes are difficult to determine. Improvement of such phenotypes therefore requires methods that can diversify large pathways and screen or select for improved variants in a quick manner. Adaptive Laboratory Evolution (ALE) provides genome-wide diversification that is usually used to study the mechanisms and changes that can drive a microbial strain to adapt under a specific pressure. However, accumulation of genomic hitchhiker mutations complicates interpretation of ALE results. Directed evolution methods restrict mutations to a defined DNA sequence, avoiding hitchhikermutations, yet most methods are unable to evolve the large sequences of DNA necessary to optimize complex phenotypes, either due to poor transformation efficiency of large plasmids or low packaging limits for M13-phage–based methods, such as PACE (Esvelt et al., 2011). Here, we introduce Inducible Directed Evolution (IDE), a method for evolving large DNA pathways using the P1 phage. IDE combines key components of previous phage-based directed evolution methods, including an inducible mutagenesis plasmid, as well as a phage that transfers mutagenized DNA to unmutated cells, producing a screenable mutant library that is often larger than can be obtained using chemical or electrical transformation methods at similar effort. Instead of the M13 phage (which has a packaging limit <5 kb), IDE employs the P1 phage, whose large packaging limit (5–100 kb) enables much larger DNA sequences to be shuttled between cells. IDE avoids the troublesome hitchhiker mutations associated with ALE and CRISPR-based methods (Wang et al., 2009; Crook et al., 2019), decouples mutagenesis and screening steps, and allows large pathways to be evolved (up to 36 kb has been published thus far). IDE’s workflow is both fast and simple to adapt. First, the diversification strain is prepared by cloning the desired DNA sequence into the phagemid backbone (see section Construction of phagemid). Once the phagemid is constructed, it is transformed to the diversification strain (see section Construction of phagemid). The diversification strain containing the P1 phage, mutagenesis plasmid, and phagemid goes through mutagenesis (see section Induction of Mutagenesis). After the mutagenesis step is completed, diversified phagemids can be produced for screening/selection in an appropriate strain (see sections Phage production, Phage infection, and Screening/selection). This cycle is described in Figure 1 and can be repeated until improvement ceases, or the desired phenotypes are obtained.
Materials and Reagents
0.22 µm filter (Genesee Scientific, catalog number: 25-227 or equivalent), store at room temperature
PCR tubes (Thermo Fisher Scientific, Eppendorf, catalog number: E0030124286 or equivalent), store at room temperature
14 mL culture tubes (Thermo Fisher Scientific, Falcon, catalog number: 14-959-1B or equivalent), store at room temperature
50 mL tubes (Thermo Fisher Scientific, Genesee Scientific Corporation, catalog number: NC1259986 or equivalent), store at room temperature
50 mL reactor tubes (Greiner Bio-One CELLSTAR® CELLreactor Conical Bottom Polypropylene Filter Top Tube 227245 or equivalent)
1.7 mL microtubes (Thermo Fisher Scientific, Genesee Scientific Corporation, catalog number: NC2045332 or equivalent), store at room temperature
Serological pipets (Thermo Fisher Scientific, Genesee Scientific Corporation, catalog number: NC0631030 or equivalent), store at room temperature
Shake flask (Thermo Fisher Scientific, PYREX, catalog number: 09-552-32 or equivalent), store at room temperature
Petri dishes (Genesee Scientific, catalog number: 32-107G or equivalent), store at room temperature
96-deep-well plates (VWR International, catalog number: 10755-248 or equivalent), store at room temperature
Pipette tips (Genesee Scientific, catalog number: 23-130RL & 24-150R & 24-160RS or equivalent), store at room temperature
Electroporation cuvette 1 mm (Genesee Scientific, catalog number: 40-100), store at room temperature
Nuclease-free water (Thermo Fisher Scientific, catalog number: J71786.K2 or equivalent), store at room temperature
Virkon (VirkonTM S Broad Spectrum Disinfectant, Chemours), store at room temperature
L-Arabinose (Teknova, catalog number: A2010 or equivalent), store at room temperature
Chloroform (Thermo Fisher Scientific, catalog number: J67241.AP or equivalent), store at room temperature
Anhydrotetracycline hydrochloride (Thermo Fisher Scientific, catalog number: J66688.MB or equivalent), store at -20 °C
Dextrose (D-Glucose) (Thermo Fisher Scientific, catalog number: D16-1 or equivalent), store at room temperature
OneTaq 2× Master Mix with Standard Buffer (New England Biolabs, catalog number: M0482S or equivalent), store at -20 °C
Q5 Site-Directed Mutagenesis Kit (New England Biolabs, catalog number: E0554S or equivalent), store at -20 °C
Q5 Hot Start High-Fidelity 2× Master Mix (New England Biolabs, catalog number: M0494S or equivalent), store at -20 °C
SGI-DNA Gibson Assembly® (GA) HiFi 1-Step Kit (CODEX DNA, Inc., catalog number: GA1200), store at -20 °C
NEBuilder® HiFi DNA Assembly Master Mix (New England Biolabs, catalog number: E2621S), store at -20 °C
NEB® 5-alpha Competent E. coli (New England Biolabs, catalog number: C2987H or equivalent). Store at -80 °C.
NEB® 10-beta Competent E. coli (New England Biolabs, catalog number: C3019H or equivalent). Store at -80 °C.
LB broth (Thermo Fisher Scientific, BD Difco, catalog number: DF0446-07-5 or equivalent), store at room temperature
Ampicillin sodium salt (Sigma-Aldrich, catalog number: A0166-25G or equivalent), store at room temperature
Chloramphenicol (VWR, catalog number: 97061-244or equivalent), store at room temperature
Kanamycin monosulfate (TCI America, catalog number: K0047 or equivalent), store at room temperature
Magnesium chloride hexahydrate (VWR, catalog number: BDH9244-500G or equivalent), store at room temperature
Calcium chloride dihydrate (Thermo Fisher Scientific, catalog number: C79-500 or equivalent), store at room temperature
TAE buffer (Tris-Acetate-EDTA) 50× (Thermo Fisher Scientific, catalog number: BP133220 or equivalent), store at room temperature
Gel loading dye (New England Biolabs, catalog number: B7024S or equivalent), store at room temperature
SYBRTM Safe DNA Gel Stain (Thermo Fisher Scientific, Invitrogen, catalog number: S33102 or equivalent), store at room temperature
Quick-Load® 1 kb Plus DNA Ladder (New England Biolabs, catalog number: N0469S or equivalent), store at room temperature
DpnI (New England Biolabs, catalog number: R0176S or equivalent), store at -20 °C
Agarose (Thermo Fisher Scientific, Lonza Inc, catalog number: BMA50004 or equivalent), store at room temperature
Plasmid Miniprep Kit (Zyppy Plasmid Miniprep Kit, Zymo Research, catalog number: D4019 or equivalent), store at room temperature
DNA Library Prep Master Mix (New England Biolabs, catalog number: E6040S or equivalent), store at -20 °C
QIAamp DNA Kit (Qiagen, catalog number: 56304), store at room temperature
DNA Clean and Concentrator-5 kit (Zymo Research, catalog number: D4004 or equivalent), store at room temperature
1× DPBS (Thermo Fisher Scientific, Gibco, catalog number: 14-190-136 or equivalent), store at room temperature
Glycerol (Thermo Fisher Scientific, catalog number: G33-4 or equivalent), store at room temperature
Sodium citrate tribasic dihydrate (Sigma-Aldrich, catalog number: C7254-1KG or equivalent), store at room temperature
Super Optimal Broth (Thermo Fisher Scientific, Bioworld, catalog number: 50-254-370 or equivalent), store at room temperature
Phagemid backbone (Addgene, catalog number: 40782)
MP6 plasmid (Addgene, catalog number: 69669)
aTc inducible promoter (Addgene, catalog number: 108529)
aTc-MP6 plasmid (Addgene, catalog number: 189935)
Primer #1: Forward primer for amplifying phagemid backbone: aggatccgaggcttggattc
Primer #2: Reverse primer for amplifying phagemid backbone: atgagatctctatgctactc
Primer #3: Mutagenic operon Forward primer: acccgtttttttggacgcgt
Primer #4: Mutagenic operon Reverse primer: agtcaaaagcctccgaccgg
Primer #5: CloDF13 Forward primer: tacgtgccaagccaaatagg
Primer #6: CloDF13 Reverse primer: ggctgacttcaggtgctaca
Primer #7: pTet and KanR Forward primer: cagtggaacgaaaaatcaat
Primer #8: pTet and KanR Reverse primer: tcagtatctctatcactgat
Primer #9: TetR Forward primer: ctcggtaccaaattccagaa
Primer #10: TetR Reverse primer: ctcacttccctgttaagtat
Note: Primers/Oligos can be ordered from Eurofins Genomics or equivalent supplier.
Ampicillin sodium salt (100 mg/mL) (see Recipes)
Chloramphenicol (34 mg/mL) (see Recipes)
Kanamycin monosulfate (50 mg/mL) (see Recipes)
10% glycerol (see Recipes)
LB media (see Recipes)
PLM (see Recipes), store at room temperature
ePLM (see Recipes), store at room temperature
Super Optimal Broth with glucose (SOC) containing 200 mM sodium citrate (see Recipes)
Equipment
iSeq 100 or equivalent (Illumina, catalog number: 20021532)
-80 °C freezer (Thermo Fisher Scientific, Revco RLE Series, model number: RLE50086A)
-20 °C freezer (Thermo Fisher Scientific, Isotemp, model number: 20LFEEFSA)
4 °C refrigerator (Jordon Refrigeration)
Microplate reader (Agilent BioTek, Synergy H1, catalog number: 11-120-535)
I26 shaking incubator (New Brunswick Scientific, Eppendorf, part number: M1324-000)
Convection incubator (VWR, Symphony, part number: 414004-610)
Incubating microplate shaker (Thermo Fisher Scientific, catalog number: 02-217-759)
Centrifuge (Thermo Fisher Scientific, Sorvall Legend X1R, catalog number: 75004261)
Microcentrifuge (Thermo Fisher Scientific, Sorvall Legend Micro 21R, catalog number: 75-002-446)
Mini Centrifuge (Thermo Fisher Scientific, catalog number: 05-090-100)
Thermocycler (Eppendorf, Mastercycler nexus eco, catalog number: 6330000021)
Gradient Thermocycler (Eppendorf, Mastercycler nexus gradient, catalog number: 6331000025)
Single channel adjustable pipettors, 0.01–1,000 µL (Eppendorf)
Multichannel adjustable pipettors 1–1,000 µL (Eppendorf)
Flow cytometer (BD Biosciences, AccuriTM C6 Plus, catalog number: 660517)
Digital dry bath heat block (Benchmark Scientific, model number: BSH1001)
MicroPulser Electroporator (Bio-Rad, catalog number: 1652100)
Spectrophotometer (Thermo Fisher Scientific, NanoDropTM 2000c, catalog number: ND-2000C)
DNA Electrophoresis Cell (Bio-Rad, Mini-Sub Cell GT Tray, catalog number: 1664400EDU)
DNA Electrophoresis Power Supply (Bio-Rad, PowerPacTM Basic, catalog number: 1645050)
Gel Imager (Bio-Rad, Gel DocTM EZ Imager, catalog number: 170-8270)
Software
Samtools (BSD, MIT, http://www.htslib.org/)
BCFtools (BSD, MIT, http://www.htslib.org/)
VarScan ( http://varscan.sourceforge.net/)
Benchling ( https://www.benchling.com) or equivalent plasmid management software
Procedure
Construction of phagemid (PM)
Use Gibson assembly (or any appropriate cloning technique) to construct the phagemid containing the pathway of interest.
The phagemid (Addgene #40782) is used as the backbone for all phagemid cloning because without an insert it turns E. coli colonies purple due to the production of violacein.
Primer #1: Forward primer for amplifying phagemid backbone: aggatccgaggcttggattc
Primer #2: Reverse primer for amplifying phagemid backbone: ctatgctactccatcgagcc
Use tmcalculator.neb.com to calculate annealing temperature.
Design primers to amplify the insert that introduce at least 20 bp homology between the fragments (see NEB Gibson Assembly protocol for instructions).
Amplify the backbone and DNA insert (20–30 ng of template) using NEB Q5 Hot Start High-Fidelity 2× Master Mix, and run the gel to check the size of the amplified fragments. Phagemid backbone amplification is 6.5kbp. Use tmcalculator.neb.com to calculate annealing temperature.
Add 1 μL of DpnI to each PCR reaction, and incubate at 37 °C for 2 h. This reaction will digest template DNA.
Purify PCR reactions using a Clean and Concentrator-5 kit according to the manufacturer’s instructions, and measure the concentration using a NanoDrop spectrophotometer.
Assemble the backbone and the insert together using NEBuilder HiFi DNA Assembly Master Mix or SGI-DNA Gibson Assembly according to the manufacturer’s instructions, except for incubating samples in a thermocycler at 50°C for 30 min instead of 15 min.
Transform the Gibson reactions to NEB 5-alpha or 10-beta Competent E. coli cells according to the manufacturer’s instructions, and grow overnight using chloramphenicol selection.
Colony PCR white colonies (purple colonies are phagemid backbone) to verify successful cloning. Colony PCR examines successful cloning before plasmid extraction. Single colonies are mixed in a PCR reaction according to themanufacturer’s instructions. The PCR reaction is prepared as the following: 0.5 µL of 10 µM forward primer, 0.5 µL of 10 µM reverse primer, 12.5 µL of OneTaq 2× Master mix, and11.5 µL nuclease-free water. Use tmcalculator.neb.com to calculate annealing temperature of the primers. Amplify junctions with NEB OneTaq Hot Start High-Fidelity 2× Master, and run PCR reactions in gel to verify amplification size. Successful amplifications are sentfor Sanger sequencing. Verified constructs with Sanger sequencing are then cultured in 2 mL of LB with appropriate antibiotics overnight in a shaking incubator (37 °C, 250 rpm). Plasmids from overnight culture wereextracted using Zyppy Plasmid Miniprep Kit according to the manufacturer’s instructions. Prepped plasmids can then be sequenced using whole plasmid sequencing services, such as plasmidsaurus (www.plasmidsaurus.com/).
Construction of Mutagenesis Plasmid (MP) –aTc-MP6 plasmid (Addgene #189935)
An Anhydrotetracycline-Inducible Mutagenesis Plasmid (aTc-MP) was constructed as follows:
Amplify the mutagenic operon (danQ926, dam, seqA, emrR, ugi, and cda1) and origin of replication (CloDF13) from the L-arabinose–inducible MP6 plasmid (Addgene #69669).
Primer #3: Mutagenic operon Forward primer: acccgtttttttggacgcgt
Primer #4: Mutagenic operon Reverse primer: agtcaaaagcctccgaccgg
Primer #5: CloDF13 Forward primer: tacgtgccaagccaaatagg
Primer #6: CloDF13 Reverse primer: ggctgacttcaggtgctaca
Amplify the anhydrotetracycline-inducible system (pTet and TetR) and kanamycin resistance gene (KanR) from pAJM.011 (Addgene #108529).
Primer #7: pTet and KanR Forward primer: cagtggaacgaaaaatcaat
Primer #8: pTet and KanR Reverse primer: tcagtatctctatcactgat
Primer #9: TetR Forward primer: ctcggtaccaaattccagaa
Primer #10: TetR Reverse primer: ctcacttccctgttaagtat
Use NEBuilder HiFi to assemble the fragments according to manufacturer’s instructions.
Transform assembly reactions to NEB 10-beta Competent E. coli cells according to the manufacturer’s instructions.
Grow transformations on LB agar plates containing kanamycin and 1% glucose. Glucose is used to repress the mutagenic operon.
Colony PCR (see section Construction of phagemid) 5–10 colonies to verify successful cloning. Amplify junctions with NEB OneTaq Hot Start High-Fidelity 2× Master, and run PCR reactions in gel to verify amplification size. Successfulamplifications are sent for Sanger sequencing. Verified constructs with Sanger sequencing are then cultured in LB containing kanamycin and 1% glucose overnight in a shaking incubator (37 °C, 250 rpm). Plasmids fromovernight culture were extracted using Zyppy Plasmid Miniprep Kit according to the manufacturer’s instructions. Prepped plasmids can then be sequenced using whole plasmid sequencing services, such as plasmidsaurus(www.plasmidsaurus.com/).
Sequence-verified aTc-MP6 can then be transformed into E. coli C600 containing the P1 phage (available upon request from Crook Lab). E. coli C600 containing both P1 phage and aTc-MP is the starting point for the construction of the diversification strain.
Preparation and transformation of electrocompetent cells to generate Diversification Strain
Inoculate the selected diversification strain (E. coli C600 strain containing mutagenesis plasmid and P1 phage) into 2 mL of LB media containing 50 µg/mL of kanamycin and grow overnight in a shaking incubator (37 °C, 250 rpm).
Dilute 500 μL of overnight culture into 50 mL of LB media (1:100 dilution), and grow in a shaking incubator (37 °C, 250 rpm) to OD600 = 0.8.
Transfer culture to a 50 mL tube and chill on ice for 10–15 min.
Spin down cells at 3,000 × g and 4 °C for 5 min.
Remove supernatant and resuspend pellets in 25 mL of ice-cold 10% glycerol.
Spin down cells at 3,000 × g and 4 °C for 5 min.
Repeat steps 5–6 two times (total 2 washes). Repeat more times if having trouble with arcing during electroporation.
Resuspend washed cells in 1 mL of 10% glycerol in microcentrifuge tubes, and spin at 3,000 × gand 4 °C for 3 min.
Resuspend pellets in 0.5 mL of 10% glycerol.
Place the electroporation cuvette on ice for 2 min.
Transfer 50 μL of the cells to a chilled microcentrifuge tube. Add 1 μL of the phagemid DNA (>50 ng).
Carefully transfer the cell/DNA mix into the chilled cuvette without introducing bubbles, and make sure that the cells deposit across the bottom of the cuvette.
Electroporate cuvette with the transformation mixture on an electroporator (e.g., Bio-Rad EC1). Time constant should be 5.0–6.0 ms for successful transformation.
Immediately add 1 mL of Super Optimal Broth (SOC) medium to the electroporation cuvette, mix by pipetting up and down, and transfer to a 1.5 mL centrifuge tube.
Place the transformation reaction in a shaking incubator (37 °C, 250 rpm) for 1 h.
Centrifuge the cells at 3,000 × g for 2 min and resuspend cells in 100 μL of DPBS.
Spread cells onto a prewarmed selective plate (e.g., LB + kanamycin + chloramphenicol), and incubate at 37 °C overnight.
Induction of mutagenesis
Subinoculate overnight culture of diversification strain in LB with 1% glucose, kanamycin, and chloramphenicol into 2 mL of LB containing kanamycin and chloramphenicol (no glucose, 1:100 dilution) in a culture tube.
Induce mutagenesis by adding 200 ng/μL of aTc (2 μL of 100 ng/μL of aTc) to the subinoculated culture.
Grow induced culture in a shaking incubator (37 °C, 250 rpm) for 8–16 h depending on desired mutation rate (3.4 × 10–7 substitutions per bp per generation).
For the first cycle of IDE, start with one colony for overnight cultures.
For cycles 2 and beyond of IDE, start with a mixed population from overnight outgrowth after infection of the diversification strain.
Phage production
Inoculate 2 μL of strains after the mutagenesis step (containing P1, aTc-MP, and the phagemid) or after the screening/selection step (containing P1 and the phagemid) into 2 mL of LB media containing chloramphenicol, andgrow overnight in a shaking incubator (37 °C, 250 rpm).
Dilute 150 μL of O/N culture into 15 mL of ePLM (1:100 dilution) in 50 mL reactor tubes, and grow in the shaking incubator (37 °C, 250 rpm) to OD600 = 0.8–1.0.
Add 150 μL of 20% L-arabinose (1/100 culture volume), and place back in the shaking incubator (37 °C, 250 rpm) for 2 h. Cultures should clarify as inFigure 2.
Transfer the cultures to a 50 mL centrifuge tube, and add 400 μL of chloroform (2.5:100 volume).
Place the tubes on ice for 5 min with gentle mixing or pipetting every 1 min.
Centrifuge at 3,000 × g and 4 °C for 10 min, and transfer the supernatant to sterile tubes for storage. Phage lysate is stable at 4 °C for 1 year and indefinitely at -80 °C.

Figure 2. Cell cultures before and after phage production. Top: Strains after the mutagenesis step or after the screening/selection step are subinoculated into fresh ePLM and grown to OD600 = 1.0. Bottom: Cell cultures are lysed after 2 h of induction with L-arabinose.Phage infection
Inoculate the screening/selection strain containing P1 into 2 mL of LB media for the screening step or LB media containing kanamycin for diversification step, and grow overnight in a shaking incubator (37 °C, 250 rpm).
Subinoculate overnight culture into ePLM (1:100 dilution), and grow in a shaking incubator (37 °C, 250 rpm) to OD600 = 1.0.
Centrifuge at 3,000 × gand 4 °C for 5 min, and resuspend the pellet in 1/3 volume of fresh ePLM.
Mix 1 mL of cells with 1 mL of phage lysate in a 14 mL culture tube.
Place the infection mixture in a standing incubator (37 °C) for 20 min and then for another 20 min in a shaking incubator (37 °C, 250 rpm).
Add 1 mL of SOC containing 200 mM of sodium citrate to the infection reaction to quench the infection. Place the reaction in a shaking incubator (37 °C, 250 rpm) for 1 h.
To eliminate the uninfected cells, transfer the infection reaction to 50 mL of LB media containing kanamycin and chloramphenicol for the diversification step or LB agar plates containing chloramphenicol for the screening step.
To count the library size of the infection, spot plate on LB agar plates containing the appropriate antibiotics.
In a 96-well plate, perform serial dilutions of the infection reaction with ePLM. Perform five dilutions, from 1:10 to 1:105. Add 180 μL of ePLM to rows A-E. In row A, add 20 μL of the infection reaction.Preform the rest of the dilutions by adding 20 μL of the previous dilution to the next row. Make sure to mix well and change pipette tips after each dilution. This step can be done with multichannel pipettors.
Using a multichannel pipettor, plate 5 μL of each infection reaction dilution on LB agar plates containing kanamycin and chloramphenicol for the diversification step or LB agar plates containing chloramphenicol forthe screening step.
Incubate spotted plates overnight in a standing incubator (37 °C).
Count CFU/mL in the morning to determine library size. Remember to multiply the number of colonies counted by the dilution factor corresponding to that row to obtain CFU/mL.
To check whether phage production and infection steps were carried correctly, run the following controls:
Infect desired strains with another phage lysate that has already generated an expected library size. This control will determine if phage production or infection failed.
Plate uninfected cells on LB agar plates containing chloramphenicol or kanamycin to test whether the infected strain has resistance to either antibiotic.
Introduction of P1 phage into P1-free strains for desired screening strains
Produce phage lysate from a strain containing P1 and phagemid.
Induce phage production (see section: Phage production).
Infect the strain of interest (see section: Phage infection).
Streak 10 μL of infection reaction on standard size LB agar plates to obtain single colonies.
Screen 10–20 colonies for P1 using P1-specific primers (fragment size 296 bp):
Forward primers: ACGACCATGAAAGCTCTTCACCCGTAG
Reverse primers: GCTTATTCGCACCTTCCCTAAAAACAAAGATGTTTGTAG
Note: To increase P1 infection efficiency to a new strain, use 1:10 cells:phage lysate by volume. Note that this ratio differs for infection with phagemid-containing particles that were discussed previously.
Screening/Selection
Freshly infected screening/selection strains are directly used from the prior step.
Screening/selection steps are different based on the application.
Example 1: Growth-based selection for increasing the utilization of a specific sugar.
Ensure that the screening/selection strain achieves a low, but nonzero growth rate in minimal media containing the sugar as the sole carbon source.
Select improved variants by culturing the library in minimal media containing the sugar as the sole carbon source.
Subinoculate culture 2–5 times to select rapidly-growing strains before transfer back to the diversification strain. Repeated subinoculations without transfer run the risk of selecting for strains withgenomic mutations.
Example 2: Biosensor-enabled screening for increasing the production of a molecule or protein.
Include a biosensor in the screening/selection strain that couples productivity to fluorescence, where wild-type productivity remains in the linear range of the biosensor.
Culture the screening/selection strain under conditions that favor biomolecule production.
Screen for high-producing variants via Fluorescence-Activated Cell Sorting (FACS).
As above, isolated variants can be grown and re-sorted via FACS several times before transfer back to the diversification strain.
Mutagenesis, Phage production, Phage infection, and Screening/Selection steps are repeated until a satisfactory variant is found.
After screening is complete
Plasmids may be extracted from screened/selected libraries and used for next-generation sequencing to measure the frequencies of common mutations. Illumina iSeq 100 was used to achieve 80–100× sequencing depth.Other sequencing machines and services that can reach similar sequencing depth can be used. Illumina whole-genome sequencing kits and IDT® for Illumina® DNA/RNA UD Indexes are used for librarypreparation and sequencing. Equivalent NGS primers can be used to index different samples. Plasmid or amplicon-based sequencing via plasmidsaurus (www.plasmidsaurus.com/) can also be used.
Trim reads using Trimmomatic.
Align trimmed reads using Bowtie2.
Identify mutations using VarScan.
A sample computational pipeline is provided below:
#!/bin/bash
java -jar VarScan.v2.3.9.jar mpileup2snp variants.pileup --min-var-freq 0.0001 > snps.txt
##trimming reads
java -jar trimmomatic-0.38.jar PE Mel1_R1.fastq R2.fastq R1_F_P.fastq R1_F_U.fastq R1_R_P.fastq R1_R_U.fastq ILLUMINACLIP:NexteraPE-PE.fa:2:30:10:2:keepBothReads LEADING:3 TRAILING:3 MINLEN:36 SLIDINGWINDOW:4:15
##Mapping to our Genome
bowtie2-build -f PM.fasta plasmidseq
bowtie2 --very-sensitive --end-to-end -x plasmidseq -1 R1_F_P.fastq -2 R1_R_P.fastq -U R1_F_U.fastq,R1_R_U.fastq -S R1.sam
samtools sort -m 10G -o R1.bam -T R1 R1.sam
samtools index -b R1.bam
samtools mpileup -f PM.fasta -d 0 R1.bam -o variants.pileup
##Calling variants
java -jar VarScan.v2.3.9.jar mpileup2indel variants.pileup --min-var-freq 0.0001 > indels.txt
Plasmids may be extracted from individual isolates and used for whole-plasmid sequencing.
Align whole-plasmid sequencing results to find mutations in isolated variants against the wild-type sequence using https://www.benchling.com or equivalent tools.
Note that the phagemid backbone may accumulate mutations during screening, potentially affecting its copy number. To directly compare evolved pathways to the wild type, evolved pathways may be cloned into a fresh phagemid backbone,or both evolved, and wild-type pathways may be integrated into the genome.
The performance of individual isolates may be compared to the screening/selection strain containing the wild-type phagemid using additional assays (e.g., growth in 96-well plates or productivity in shake flasks, followed byHPLC analysis).
The most productive variants may be used as is or subjected to additional rounds of IDE.
Appendix 1: Running gels
Confirm cloning and PCR amplifications using a 1% agarose gel.
Dissolve 300 mg of agarose in 30 mL of 1× TAE buffer by heating until boiling.
Add 3 μL of SYBR Safe DNA gel stain to the liquid gel.
Cast the gel by pouring into gel trays with combs in place and allow to solidify.
Run the loaded gel with 5 μL of Quick-Load 1 kb Plus DNA Ladder at 120 V for 25–30 min. Power is supplied by BioRad PowerPac, and the gel is run in a BioRad DNA electrophoresis chamber.
Image the gel using a Gel Doc EZ Imager or equivalent.
Appendix 2: Best practices for handling phage:
All steps containing phage lysate should be conducted in the BSL2 hood to avoid contaminating other E. coli experiments. Quarantine your P1 work as much as possible from your other projects. This can be done by designating a separate bench space and materials for projects involving phages.
Use filter tips when pipetting P1 phage lysate.
Use Virkon to wipe off all surfaces and tools after handling phages.
Store phage lysate in screw-top 15 or 50 mL centrifuge tubes. Infections and phage production are also done in these tubes.
If possible, store P1 lysates in a 4 °C fridge that is not used for other E. coli experiments.
Do not keep old plates of bacteria containing P1 as phages can initiate lysis in stationary colonies (<1 week in 4 °C).
Recipes
Ampicillin sodium salt (100 mg/mL)
Dissolve 5 g of ampicillin into a 50 mL Falcon tube of ddH2O, and filter through a 0.22 µm filter to sterilize. Aliquot 1 mL and store at -20 °C (use at 1:1000 dilution in LB or LB-Agar).
Chloramphenicol (34 mg/mL)
Dissolve 1.7 g of chloramphenicol to 30 mL of 100% ethanol in a 50 mL Falcon tube, and bring volume to 50 mL with 100% ethanol. Aliquot 1 mL into microcentrifuge tubes, and store at -20 °C (use at 1:1,000 dilution in LB or LB-Agar).
Kanamycin monosulfate (50 mg/mL)
Add 2.5 g of kanamycin monosulfate to 30 mL of DI water in a 50 mL Falcon tube, and then bring volume to 50 mL with DI water. Filter through a 0.22 µm filter to sterilize. Aliquot 1 mL into microcentrifuge tubes, and store at-20 °C (use at 1:1,000 dilution in LB or LB-Agar).
10% glycerol
Reagent Final concentration Amount Glycerol 10% 10 mL Nuclease-free H2O n/a 90 mL Total n/a 100 mL LB media
Dissolve 25 g/L of LB broth in DI water, and autoclave the final solution at 121 °C for 20 min. Tighten the caps, and store at room temperature once fully cooled.
PLM
Autoclave the final solution at 121 °C for 20 min.
Reagent Final concentration Amount LB broth n/a 25 g MgCl·6HO 100 mM 20.3 g CaCl2·2H2O 5 mM 0.7 g H2O n/a 100 mL Total n/a 100 mL ePLM
Autoclave the final solution at 121 °C for 20 min.
Reagent Final concentration Amount LB broth n/a 25 g MgCl·6HO 140 mM 28.4 g CaCl2·2H2O 7 mM 1 g H2O n/a 100 mL Total n/a 100 mL Super Optimal Broth with glucose (SOC) containing 200 mM of sodium citrate
Reagent Final concentration Amount Super Optimal Broth n/a 28 g Sodium citrate tribasic dihydrate 200 mM 58.8 g H2O n/a 1 L Total n/a 1 L Autoclave once dissolved. Allow the solution to cool to 50 °C or below before adding 20 mL of 1 M MgSO4 and 20 mL of 1 M glucose. Store at room temperature.
Acknowledgments
We thank the labs of Dr Christopher Anderson (UC Berkeley) for phagemid constructs (Addgene #40782), Dr. David R. Liu (Harvard University) for the MP6 plasmid (Addgene #69669), and Dr. Chase Beisel for wild-type and engineered P1 bacteriophages.
This protocol has been adapted from Al’Abri et al. (2022).
Competing interests
I.S.A. and N.C. have a patent pending related to this work.
References
- Al'Abri, I. S., Haller, D. J., Li, Z. and Crook, N. (2022). Inducible directed evolution of complex phenotypes in bacteria. Nucleic Acids Res 50(10): e58.
- Crook, N., Ferreiro, A., Gasparrini, A. J., Pesesky, M. W., Gibson, M. K., Wang, B., Sun, X., Condiotte, Z., Dobrowolski, S., Peterson, D., et al. (2019). Adaptive Strategies of the Candidate Probiotic E. coli Nissle in the Mammalian Gut. Cell Host Microbe 25(4): 499-512.
- Esvelt, K. M., Carlson, J. C. and Liu, D. R. (2011). A system for the continuous directed evolution of biomolecules. Nature 472(7344): 499-503.
- Wang, H. H., Isaacs, F. J., Carr, P. A., Sun, Z. Z., Xu, G., Forest, C. R. and Church, G. M. (2009). Programming cells by multiplex genome engineering and accelerated evolution. Nature 460(7257): 894-898.
Article Information
Copyright
© 2022 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Al'Abri, I. S., Li, Z., Haller, D. J. and Crook, N. (2022). A Novel Method of Inducible Directed Evolution to Evolve Complex Phenotypes. Bio-protocol 12(20): e4535. DOI: 10.21769/BioProtoc.4535.
Category
Biological Engineering > Synthetic biology
Molecular Biology > DNA > Mutagenesis
Molecular Biology > DNA > Transfection
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.