- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Isolation and Characterization of Membrane Vesicles from Lactobacillus Species
Published: Vol 11, Iss 17, Sep 5, 2021 DOI: 10.21769/BioProtoc.4145 Views: 5235
Reviewed by: Alba BlesaJing LiChenchen Liu
Original research article
The authors used this protocol in:

Jan 2019
Advertisement



Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Related protocols
Abstract
Throughout their life cycle, bacteria shed portions of their outermost membrane comprised of proteins, lipids, and a diversity of other biomolecules. These biological nanoparticles have been shown to have a range of highly diverse biological activities, including pathogenesis, community regulation, and cellular defense (among others). In recent publications, we have isolated and characterized membrane vesicles (MVs) from several species of Lactobacilli, microbes classified as commensals within the human gut microbiome (Dean et al., 2019 and 2020). With increasing scientific understanding of host-microbe interactions, the gut-brain axis, and tailored probiotics for therapeutic or performance increasing applications, the protocols described herein will be useful to researchers developing new strategies for gut community engineering or the targeted delivery of bio-active molecules.
Graphic abstract:
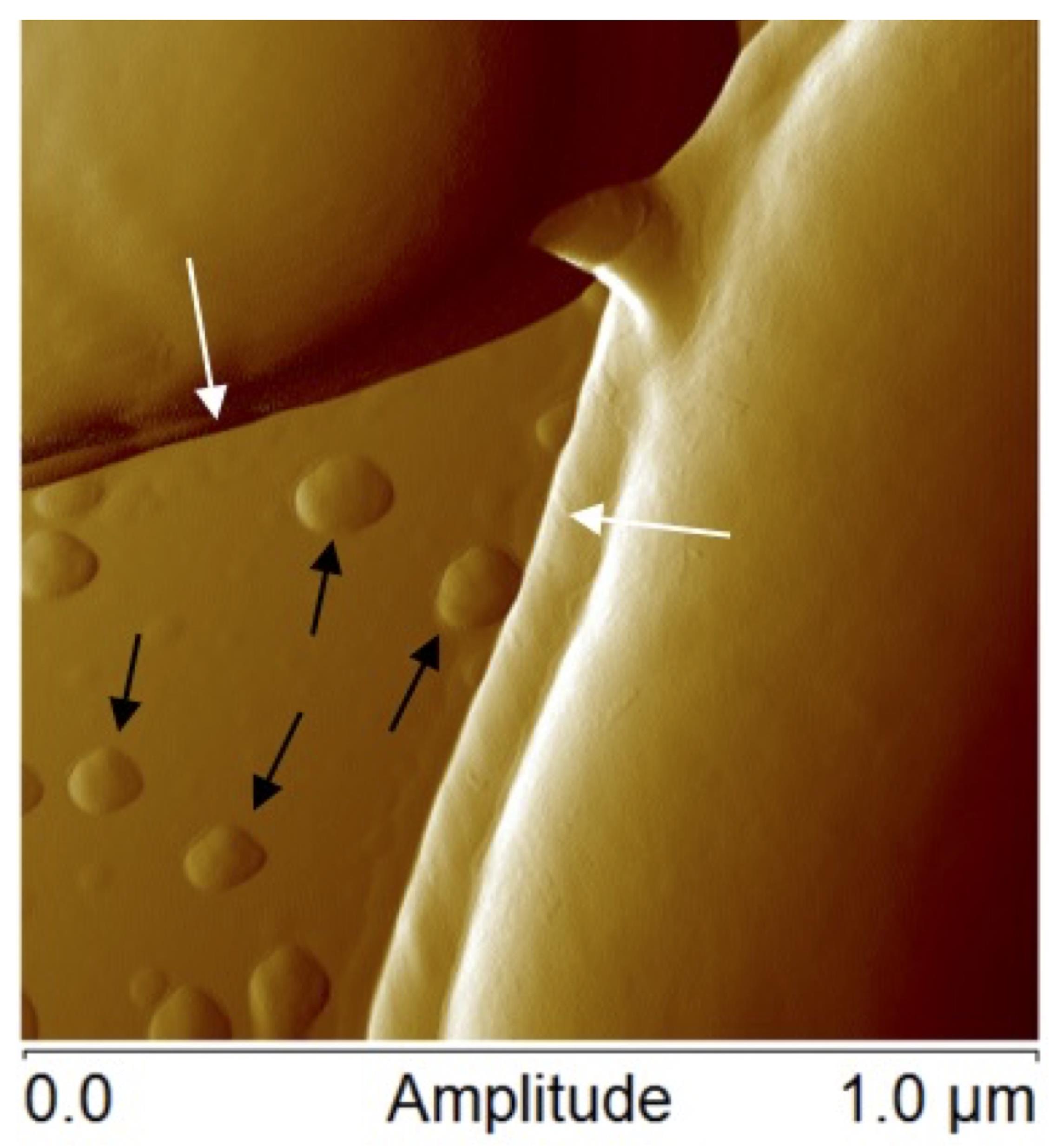
Figure 1. Atomic force microscopic image of Lactobacillus casei ATCC 393 bacteria margins (white arrows) and membrane vesicles (black arrows)
Background
The membranes of all cells are dynamic structures, the biomolecule composition of which is constantly changing as cells respond to environmental stimuli, alter protein and lipid composition, release waste products, take in nutrients, and perform many other cellular processes (Vereb et al., 2003; Watson, 2015). Throughout cellular life cycles, fragments of the outermost membrane are often shed as nanosized particles. In bacteria, these structures are often referred to as membrane vesicles (MVs) or outer membrane vesicles (OMVs), which will be referred to as MVs throughout for simplicity (a representative example of MVs from Lactobacillus casei is shown in Figure 1). As interest in these biological nanoparticles has grown in recent years, researchers have shown that MVs have broad biological activities, including host-microbe interactions, gene transfer, and community regulation (Kulp and Kuehn, 2010; Caruana and Walper, 2020).
Bacterial MVs have shown significant promise for applications such as vaccine development and potential therapeutic applications. Naturally occurring OMVs from Gram-negative bacteria have shown significant promise in the development of vaccines for bacteria such as Neisseria meningitidis and Burkholderia pseudomallei, pathogens that have proven challenging to vaccine and therapeutic development alike (Holst et al., 2009; Nieves et al., 2014). Recently, research groups have also shown that the MVs from some commensal bacteria can also modulate responses from host immune systems and stimulate host neurological systems (Mata Forsberg et al., 2019; Molina-Tijeras et al., 2019; Rodovalho et al., 2020). With growing capabilities in synthetic biology, the potential uses of MVs are steadily increasing as researchers have developed engineering strategies that allow for modification of genetic and cellular pathways to control the composition of nascent MVs. These efforts have led to new biomaterials for therapeutic applications, environmental decontamination, and other purposes (Alves et al., 2018; Qing et al., 2019).
The classification of lactic acid bacteria (LAB) encompasses several genera of bacteria with similar characteristics of acid-tolerance and fermentation capabilities. Many LAB are classified as generally regarded as safe (GRAS) and have been used in the production of food products for centuries. Additionally, several LAB have been recognized as beneficial to their host and are studied for their health benefits as probiotics leading to a large commercial market for probiotic supplements and foods. While live-bacterial cultures are the most commonly used form of probiotics, the potential for engineering or enriching for specific cellular components has led researchers to explore the use of purified MVs for controlled therapeutic applications (Seo et al., 2018; Molina-Tijeras et al., 2019; Dean et al., 2020).
There are numerous protocols for the purification of MVs from eukaryotic cells and Gram-negative bacteria, which have been the primary focus of MV research (Klimentova and Stulik 2015; Alves et al., 2017; Dauros Singorenko et al., 2017). Recently, there has been growing interest in the MVs of gut bacteria and the roles they may play in host and community interactions. LAB are Gram-positive bacteria and therefore have a significantly different membrane and peptidoglycan structure as compared to Gram-negative bacteria. While this may not specifically contribute to biophysical differences between Gram-negative and Gram-positive MVs, it has been shown that the MVs of some LAB species have a bimodal size distribution with an abundant population of smaller MVs in the 10-50 nm size range (Dean et al., 2019). Here, we describe protocols for the isolation and characterization of MVs that have proven successful for numerous LAB species. MVs are isolated from overnight LAB cultures via ultracentrifugation and then analyzed for concentration and relative size distribution using a NanoSight nanoparticle tracking instrument. Dynamic light scattering (DLS) is used as another way to measure size distribution and also to measure zeta potential (surface charge). Then, the protein content of MVs can be examined both qualitatively using SDS-PAGE and specifically by using mass spectrometry for proteomic analysis (schematic shown in Figure 2). While this manuscript describes work we have employed for LAB, these protocols could also be used for the isolation of MVs from a variety of microbial species.
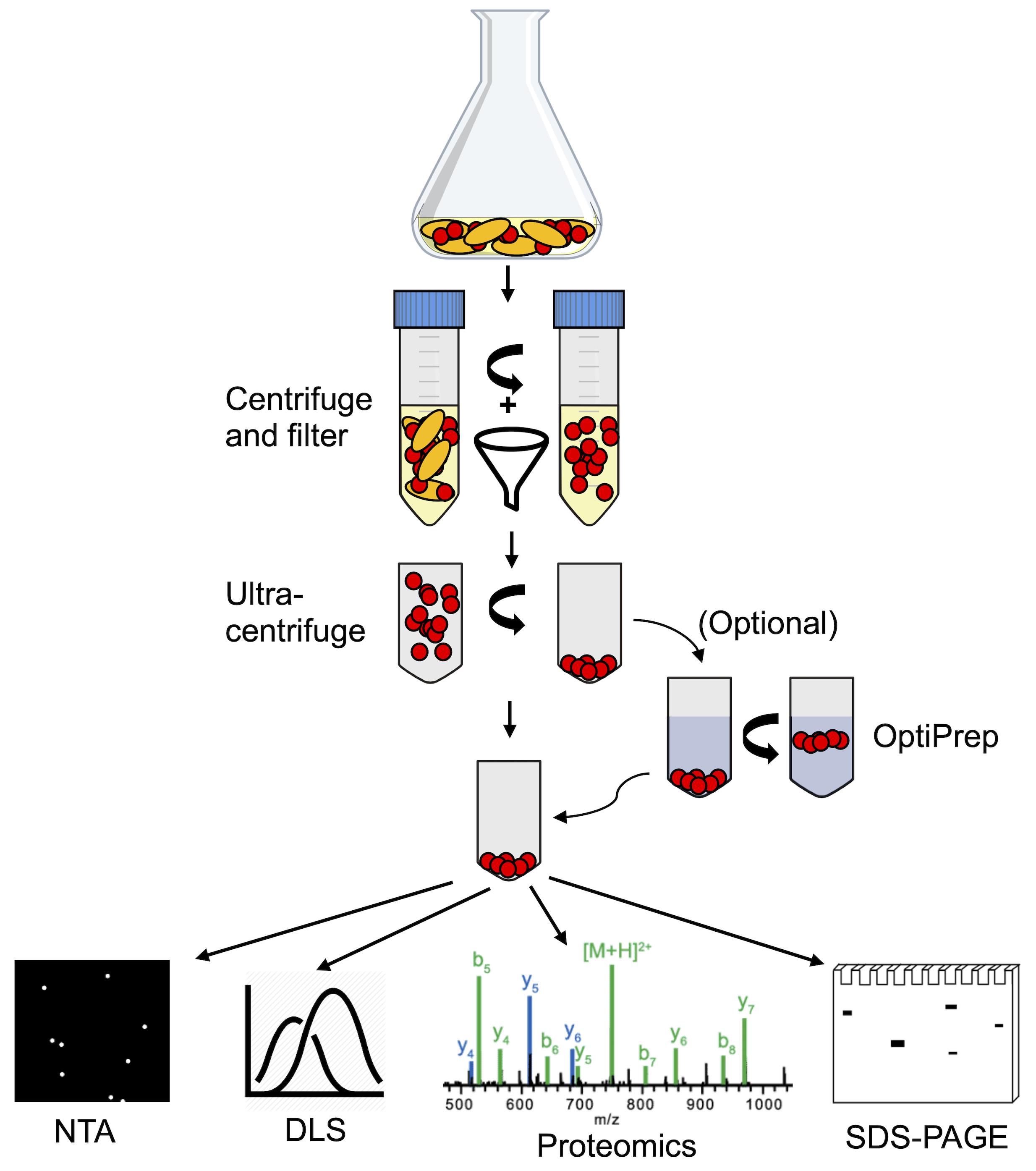
Figure 2. Schematic overview of the membrane vesicle isolation and characterization process. MVs are purified from Lactobacillus cultures initially via centrifugation and filtration, removing cells and large cellular debris. From the filtered supernatant, MVs are isolated by ultracentrifugation. Gradient centrifugation using OptiPrep medium can be applied to isolated MVs as an additional purification step. Purified MVs can then be characterized by nanoparticle tracking analysis (NTA), dynamic light scattering (DLS), SDS-PAGE protein gels, shotgun proteomics, or other methods.
Materials and Reagents
AnaeroJarTM 2.5 L jars (Oxoid®, ThermoFisher Scientific, catalog number: AG0025A)
AnaeroGenTM anaerobic gas generation sachets (Oxoid®, Thermo Fisher Scientific, catalog number: AN0025A)
Resazurin anaerobic indicator strips (Oxoid®, Thermo Fisher Scientific, catalog number: BR0055B)
100 mm sterile Petri dishes (Fisherbrand, Thermo Fisher Scientific, catalog number FB0875712)
Pyrex® Erlenmyer flasks 250 ml (CorningTM, Thermo Fisher Scientific, catalog number: CLS4980250)
50 ml conical centrifuge tubes (CorningTM Falcon, Thermo Fisher Scientific, catalog number: 14-432-22)
0.45 µm syringe filters (J. G. Finneran, catalog number: FEC0425PC)
30 ml sterile syringes (BD Slip tip sterile syringe; Thermo Fisher Scientific, catalog number: BD 302833)
38.5 ml ultracentrifuge tubes (Thinwall Ultra-Clear tubes; Beckman Coulter, catalog number: 344058)
5 ml ultracentrifuge tubes (Thinwall Ultra-Clear tubes; Beckman Coulter, catalog number: 344057)
1 ml needle-less syringes (Henke Sass Wolf, catalog number: 4010.200V0)
1.5 ml centrifuge tubes (Thermo Fisher Scientific, catalog number: P190411)
Disposable cells compatible with ZetaSizer Nanoseries (Malvern Panalytical, catalog number: DTS 1070)
Lactobacillus species, i.e.,
Lactobacillus acidophilus (ATCC 53544)
Lactobacillus casei (ATCC 393)
Lactobacillus reuteri (ATCC 23272)
Lactobacillus plantarum (ATCC BAA-793)
de Man, Rogosa, and Sharpe (MRS) media (Sigma-Aldrich, catalog number: 69966-500G, prepare according to the manufacturer’s instructions)
Tween® 80 (Sigma Aldrich, catalog number: P8074)
4-15% Mini-PROTEAN® TGX Pre-cast protein gels (Bio-Rad, catalog number:4561085)
GelCodeTM Blue Stain Reagent (Thermo Fisher Scientific, catalog number: PI24590)
10× phosphate-buffered saline (Thermo Fisher Scientific, catalog number: AM9625)
4× Laemmli sample buffer (Bio-Rad, catalog number: 1610747)
2-mercaptoethanol (Bio-Rad, catalog number: 161-0710)
SDS-PAGE running buffer (Bio-Rad, catalog number: 161-0772)
Pierce BCA Protein Assay Kit (Thermo Fisher Scientific, catalog number: 23225)
1-propanol (Sigma-Aldrich, catalog number: 402893)
Ammonium bicarbonate (Sigma-Aldrich, catalog number: A6141)
Trypsin, sequencing grade (Promega, Fisher Scientific, catalog number: PRV5111)
Formic acid (Sigma-Aldrich, catalog number: 27001)
Acetonitrile (Sigma-Aldrich, catalog number: 34851)
OptiPrep medium (Progen Biotechnik GmbH, catalog number: 1114542)
SDS-PAGE Running Buffer (10× stock; use at 1×) (see Recipes)
Equipment
Incubator capable of maintaining 37°C (for example: Fisherbrand, Isotemp Microbiological Incubator)
Centrifuge with capacity for 50 ml conical tubes (for example: Beckman Coulter, Avanti JXN-30 using a JA-14.50 rotor)
Ultracentrifuge with capacity for 38.5 ml tubes, capable of 129,000 × g (for example: Sorvall WX Ultra 90 centrifuge using AH-629 rotor)
NanoSight LM10 (Malvern Panalytical, Worcestershire, UK)
ZetaSizer NanoSeries equipped with a HeNe laser source (λ = 633 nm) and avalanche photodiode (Malvern Panalytical, Worcestershire, UK)
Mini-PROTEIN® Tetra Vertical Electrophoresis Cell (Bio-Rad, catalog number: 1658004)
Speed-vac (for example: Thermo Fisher Scientific SC210A SpeedVac Concentrator, catalog number: SC210A-230)
Barocycler (Pressure Biosciences Inc., HUB 440-SW16, Easton, MA, US)
Orbitrap LC-MS/MS system (for example: Thermo Scientific Orbitrap Fusion Lumos equipped with a Nanospray Flex Ion Source (Thermo Fisher Scientific, catalog number: ES071)
Autosampler (for example: Thermo Scientific Dionex UltiMate 3000 Rapid Separation Well Plate Autosampler (Thermo Fisher Scientific, catalog number: 5840.0010)
Ultra-high performance liquid chromatography (UHPLC) system (for example: Dionex Ultimate 3000 RSLCnano system, Thermo Fisher Scientific, catalog number: ULTIM3000RSLCNANO)
Software
NTA 2.3 Nanoparticle Tracking and Analysis software (Malvern Panalytical, Worcestershire, UK)
Dispersion Technology Software (DTS, Malvern Panalytical, Worcestershire, UK) used for dynamic light scattering analysis
Scaffold version 4.8.2 (Proteome Science Inc., Portland, Oregon, US), used for protein identification after mass spectrometry
Mascot (version 2.6.1, Matrix Science, London, UK), used for protein identification after mass spectrometry
X! Tandem (version 1.7.18) used for protein identification after mass spectrometry
R (https://cran.r-project.org/) and relevant R packages: Peptides, Limma, and ggplot2. Used for biochemical analysis of identified proteins, statistical analysis, and generation of volcano plots or other visualizations of the data
Procedure
Isolation of membrane vesicles (MVs)
Grow bacterial strains
Streak each strain individually from frozen stocks or stab vials onto MRS agar plates. For anaerobic growth, place plates in an AnaeroJar jar with an AnaeroGen anaerobic gas generation sachet and anaerobic indicator added. Grow at 37°C for approximately 48 h or until individual colonies are visible.
Inoculate a single bacterial colony into 3 ml MRS broth. Grow overnight at 37°C without shaking in an AnaeroJar as in the previous step.
Add 1 ml from the primary culture to 50 ml MRS broth in a sterile 250 ml Erlenmeyer flask. Grow overnight as in previous steps.
Collect and filter supernatant
Transfer the overnight culture into a 50 ml conical centrifuge tube and centrifuge at 5,000 × g for 30 min.
Decant supernatant into a new tube and centrifuge again at 5,000 × g for 30 min.
Repeat Step A2b once or twice more, until no cell pellet is visible on the sides or bottom of the tube after centrifugation. These additional centrifugation steps are not strictly required but make it easier to filter the supernatant in the following step.
Pass the supernatant through a 0.45 µm filter. This may be done using a 30 ml syringe and syringe filter or using a vacuum apparatus with an appropriate filter for larger volumes of supernatant.
Ultracentrifugation to isolate MVs
Transfer 36 ml of the filtered supernatant to an ultracentrifuge tube and assemble into the rotor.
Ultracentrifuge at 126,000 × g for 1.5 h at 4°C.
Decant the supernatant (the pellet is often invisible).
Add 1 ml of 1× phosphate-buffered saline (PBS) and incubate overnight at 4°C to resuspend the pellet.
Note: If proceeding for gradient ultracentrifugation, resuspend in 150 μl of PBS-OptiPrep mixture instead (see Step A4a below).
Gradient ultracentrifugation of MVs
Resuspend MV pellets in 150 µl of a PBS-OptiPrep mixture (45% v/v) and move the suspension to the bottom of a 5 ml ultracentrifuge tube.
Make several 0.8 ml PBS-OptiPrep solutions of lower density (e.g., 35%, 30%, 25%, 20%, 15%, and 10%) and sequentially cover the base layer containing MVs with layers of decreasing density. Avoid adding bubbles or disrupting previous layers as you add.
After all layers have been added, allow the tube to settle for 1 h.
Ultracentrifuge the gradient at 236,000 × g for 3 h in a Sorvall WX Ultra 90 centrifuge using an AH-650 rotor.
Following centrifugation, gently sequentially remove and place 10 equal-volume (~500 μl) fractions into separate tubes.
Assess the different density fractions for MV concentration, protein composition, and other characteristics using the methods described below.
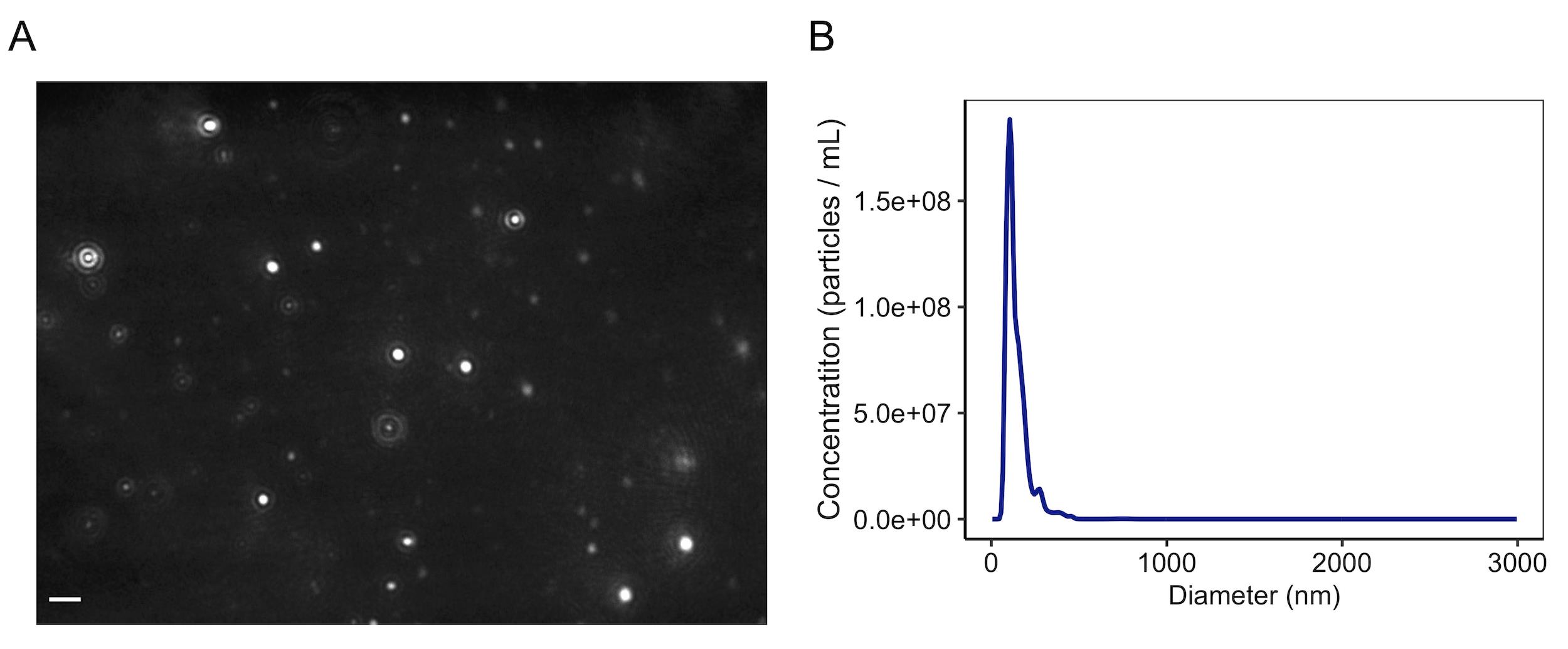
Figure 3. Nanoparticle tracking analysis (NTA) of Lactobacillus membrane vesicles. A. Representative frame taken from a NanoSight LM10 video of L. acidophilus MVs. MVs purified from an MRS culture were diluted 1:1,000 in PBS prior to NTA. Scale bar indicates 2 µm. B. Representative MV size distribution from NTA results, taken from an average obtained from three 60 s videos.
MV characterization
Nanoparticle tracking analysis (NTA) to determine particle size distribution and concentration
Dilute MVs in 1× PBS – make several serial dilutions such as 10×, 100×, and 1,000×. Detection of particles by the NTA software is best when there are 20-60 particles within the field of view; therefore, samples need to be diluted, and the optimal dilution for each sample may vary from one isolation to another.
Using a 1 ml needle-less syringe, add the diluted sample to the NanoSight instrument chamber. Allow a moment for samples to settle and adjust microscope focus and camera settings to optimize clarity of particles in the camera field of view within the NTA 2.3 Nanoparticle Tracking and Analysis software. Data can be collected at room temperature.
Capture data as triplicate readings of 60 s exposure at 30 frames per second, and analyze using auto particle detection and tracking parameters: detection threshold, pixel blur, minimum track length, and minimum expected particle size. A representative image of NTA of vesicles from Lactobacillus acidophilus ATCC 53544 is shown in Figure 3. As NTA 2.3 does not automatically add scale bars to images, they can be added by converting pixels to µm by dividing pixels by 9.036 (when using a 30× objective).
Dynamic light scattering (DLS) to measure MV size distribution
Dilute MV samples 10-fold in 0.1× PBS pH 7.4 and add to disposable DLS cuvettes.
Carry out DLS analysis using ZetaSizer NanoSeries equipped with a HeNe laser source (λ = 633 nm) and analyze using Dispersion Technology Software. Carry out measurements at 25°C. For each sample, take the average of five runs of 10 s each for the autocorrelation function, and repeat this three to six times.
Zeta Potential measurement to determine MV surface charge
Dilute MV samples and load into disposable cuvettes as for DLS.
Measure zeta potential using ZetaSizer NanoSeries equipped with a HeNe laser source (λ = 633 nm) with Dispersion Technology Software. Carry out measurements at 25°C in triplicate for each sample.
SDS-PAGE for qualitative analysis of proteins contained in MVs
Prepare samples: based on MV concentration as determined by NTA, take equal amounts of each MV sample in ≤15 μl volume in 1.5 ml centrifuge tubes. Adjust sample volumes to 15 μl using water or 1× PBS. (Volumes may be adjusted depending on the well capacity of SDS-PAGE gels to be used.)
Add 5 μl (or required volume to result in 1× final concentration) of 4× Laemmli sample loading buffer to each sample, then denature sample proteins by boiling for 5 min, chilling on ice for 5 min, and centrifuging for 15 min at 16,000 × g in a benchtop centrifuge.
Assemble a Bio-Rad Mini-PROTEAN Vertical Electrophoresis cell according to the manufacturer’s instructions with a 4-15% Mini-PROTEAN® TGX pre-cast protein gel.
Load denatured MV samples (supernatant only) into the gel and run at 200 V until the dye front reaches the bottom of the gel.
Remove the gel from its casing and rinse briefly in water; then, stain in GelCodeTM Blue Stain Reagent, followed by destaining using water, to visualize MV protein bands. Figure 3 of Dean et al. (2019) may be referred to for an example of an SDS-PAGE gel of MVs from three LAB species.
Shotgun Proteomics Analysis to identify protein contents of MVs
Sample preparation
Resuspend MV pellets with 10 ml of 10% n-propanol in 50 mM ammonium bicarbonate (ABC) buffer.
Normalize sample protein concentration using the Pierce BCA Protein Assay Kit.
Digest samples (50 µg) in solution with sequencing-grade modified trypsin at a 1:50 (w/w) enzyme:substrate ratio in a barocycler for 90 min (90 cycles: 50 s on at 45 kpsi, 10 s off).
Evaporate digested samples via speed-vac and store at -20°C until analysis by LC-MS/MS.
LC-MS/MS
Reconstitute dried samples in 0.1% formic acid in water (solvent A) and load 3 μg digested protein onto a reversed-phase C18 column by a Dionex Ultimate WPS-3000 autosampler connected to an Orbitrap Fusion Lumos equipped with a Nanospray Flex Ion Source in data-dependent acquisition mode.
Separate peptides across a 90 min gradient of 2-60% solvent B (0.1% formic acid in acetonitrile) at a flow rate of 300 nl/min using a Dionex Ultimate 3000 RSLCnano system.
Save the raw, mgf, and mzid files output for each sample for downstream analysis.
Protein identification
Search mass spectrometry data using Mascot and X! Tandem using the appropriate genome file for the organism. Validate peptide-spectrum matches with Scaffold.
Set fragment ion mass tolerance to ±0.60 Da and parent ion tolerance to ±0.60 Da. Set proteins identification thresholds to ≥2 peptides (protein probability 80%, peptide probability 95%). Export list of identified proteins to CSV.
Analyze the output CSV files with R.
Data analysis
Proteomics analysis:
Analyze all MS/MS samples using Mascot (Matrix Science, London, UK) and X! Tandem (The GPM, thegpm.org). Have Mascot set up to search the correct database sequence file (generated by protein .fasta input) and a corresponding common contaminants file. Set assumed cleavages to digestion using trypsin.
Set up X! Tandem to search a reverse concatenated subset of the correct database sequence file, which only contains proteins identified by Mascot in analyzed samples.
Use Scaffold (Proteome Software Inc., Portland, OR, USA) to validate MS/MS based peptide and protein identifications. Set peptide-spectrum match settings to: fragment ion mass tolerance = ±0.60 Da and the parent ion tolerance = ±0.60 Da. Set protein identification thresholds to ≥2 peptides (protein probability 80%; peptide probability 95%).
Analyze proteomics data output to CSV files with R
Determine physicochemical characteristics of peptides/proteins using the Peptides R package.
Perform the two-sample t-tests using an empirical Bayes method to adjust the estimate of variance of each protein using the Limma R package, the output of which is each protein with its log2 fold-change and moderated p-value corresponding to the moderated t-statistic, as previously described (Dean et al., 2020).
Use the ggplot2 R package to create volcano plots and other visualizations of the data (see Figure 4 for an example volcano plot using proteomics data).
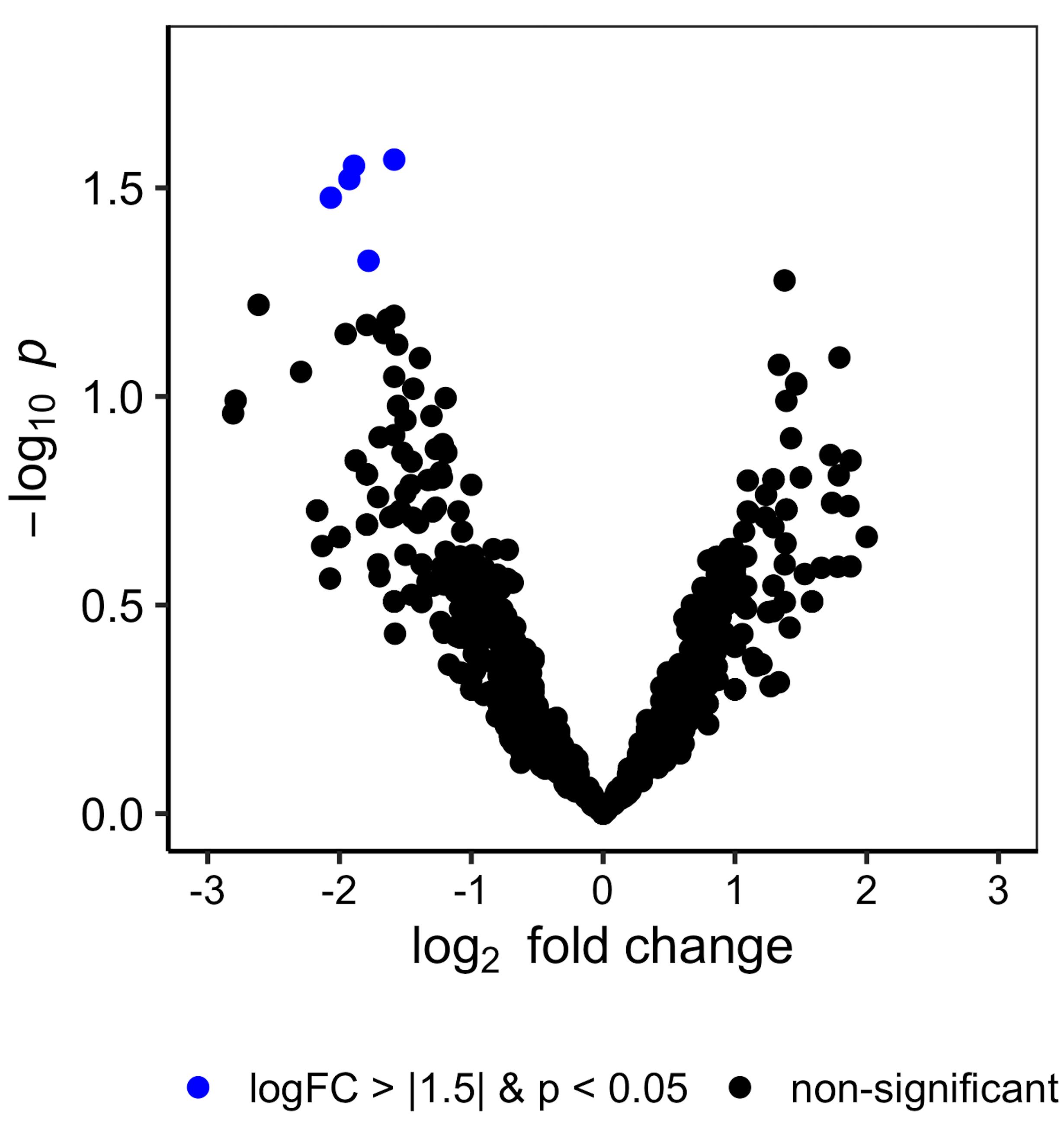
Figure 4. Volcano plot of proteomics performed on Lactobacillus membrane vesicles. Representative volcano plot of log2 fold change of proteins identified in L. acidophilus MVs treated with inducer relative to an untreated control. Identified protein data in triplicate samples were put through the empirical Bayes method based on the Limma R package and plotted using ggplot2. Proteins with significant change are colored in blue.
Recipes
SDS-PAGE Running Buffer (10× stock; use at 1×)
25 mM Tris base
192 mM glycine
0.1% SDS
Dissolve in water as solvent
Acknowledgments
This protocol is adapted from previous work (Dean et al., 2019 and 2020). Funding for this effort was provided through Core funds of the Naval Research Laboratory (SND, SAW). Additional financial support was provided through the American Society for Engineering Education (ASEE) post-doctoral fellowship program (JCC). The AFM image shown in the graphical abstract was taken by Claretta Sullivan, Air Force Research Laboratory, Wright Patterson Air Force Base.
Competing interests
The authors declare no competing interests.
Ethics
None of the protocols described here include studies with animals or human subjects.
References
- Alves, N. J., Moore, M., Johnson, B. J., Dean, S. N., Turner, K. B., Medintz, I. L. and Walper, S. A. (2018). Environmental Decontamination of a Chemical Warfare Simulant Utilizing a Membrane Vesicle-Encapsulated Phosphotriesterase. ACS Appl Mater Interfaces 10(18): 15712-15719.
- Alves, N. J., Turner, K. B., DiVito, K. A., Daniele, M. A. and Walper, S. A. (2017). Affinity purification of bacterial outer membrane vesicles (OMVs) utilizing a His-tag mutant. Res Microbiol 168(2): 139-146.
- Caruana, J. C. and Walper, S. A. (2020). Bacterial Membrane Vesicles as Mediators of Microbe - Microbe and Microbe - Host Community Interactions. Front Microbiol 11: 432.
- Dauros Singorenko, P., Chang, V., Whitcombe, A., Simonov, D., Hong, J., Phillips, A., Swift, S. and Blenkiron, C. (2017). Isolation of membrane vesicles from prokaryotes: a technical and biological comparison reveals heterogeneity. J Extracell Vesicles 6(1): 1324731.
- Dean, S. N., Leary, D. H., Sullivan, C. J., Oh, E. and Walper, S. A. (2019). Isolation and characterization of Lactobacillus-derived membrane vesicles. Sci Rep 9(1): 877.
- Dean, S. N., Rimmer, M. A., Turner, K. B., Phillips, D. A., Caruana, J. C., Hervey, W. J. t., Leary, D. H. and Walper, S. A. (2020). Lactobacillus acidophilus Membrane Vesicles as a Vehicle of Bacteriocin Delivery. Front Microbiol 11: 710.
- Holst, J., Martin, D., Arnold, R., Huergo, C. C., Oster, P., O'Hallahan, J. and Rosenqvist, E. (2009). Properties and clinical performance of vaccines containing outer membrane vesicles from Neisseria meningitidis. Vaccine 27 Suppl 2: B3-12.
- Klimentova, J. and Stulik, J. (2015). Methods of isolation and purification of outer membrane vesicles from gram-negative bacteria. Microbiol Res 170: 1-9.
- Kulp, A. and Kuehn, M. J. (2010). Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu Rev Microbiol 64: 163-184.
- Mata Forsberg, M., Bjorkander, S., Pang, Y., Lundqvist, L., Ndi, M., Ott, M., Escriba, I. B., Jaeger, M. C., Roos, S. and Sverremark-Ekstrom, E. (2019). Extracellular Membrane Vesicles from Lactobacilli Dampen IFN-γ Responses in a Monocyte-Dependent Manner. Sci Rep 9(1): 17109.
- Molina-Tijeras, J. A., Galvez, J. and Rodriguez-Cabezas, M. E. (2019). The Immunomodulatory Properties of Extracellular Vesicles Derived from Probiotics: A Novel Approach for the Management of Gastrointestinal Diseases. Nutrients 11(5): 1038.
- Nieves, W., Petersen, H., Judy, B. M., Blumentritt, C. A., Russell-Lodrigue, K., Roy, C. J., Torres, A. G. and Morici, L. A. (2014). A Burkholderia pseudomallei outer membrane vesicle vaccine provides protection against lethal sepsis. Clin Vaccine Immunol 21(5): 747-754.
- Qing, G., Gong, N., Chen, X., Chen, J., Zhang, H., Wang, Y., Wang, R., Zhang, S., Zhang, Z., Zhao, X., et al. (2019). Natural and engineered bacterial outer membrane vesicles. Biophysics Reports 5(4): 184-198.
- Rodovalho, V. R., da Luz, B. S. R., Rabah, H., do Carmo, F. L. R., Folador, E. L., Nicolas, A., Jardin, J., Briard-Bion, V., Blottiere, H., Lapaque, N., Jan, G., Le Loir, Y., de Carvalho Azevedo, V. A. and Guedon, E. (2020). Extracellular Vesicles Produced by the Probiotic Propionibacterium freudenreichii CIRM-BIA 129 Mitigate Inflammation by Modulating the NF-κB Pathway. Front Microbiol 11: 1544.
- Seo, M. K., Park, E. J., Ko, S. Y., Choi, E. W. and Kim, S. (2018). Therapeutic effects of kefir grain Lactobacillus-derived extracellular vesicles in mice with 2,4,6-trinitrobenzene sulfonic acid-induced inflammatory bowel disease. J Dairy Sci 101(10): 8662-8671.
- Vereb, G., Szollosi, J., Matko, J., Nagy, P., Farkas, T., Vigh, L., Matyus, L., Waldmann, T. A. and Damjanovich, S. (2003). Dynamic, yet structured: The cell membrane three decades after the Singer-Nicolson model. Proc Natl Acad Sci U S A 100(14): 8053-8058.
- Watson, H. (2015). Biological membranes. Essays Biochem 59: 43-69.
Article Information
Copyright
© 2021 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Caruana, J. C., Dean, S. N. and Walper, S. A. (2021). Isolation and Characterization of Membrane Vesicles from Lactobacillus Species. Bio-protocol 11(17): e4145. DOI: 10.21769/BioProtoc.4145.
Category
Microbiology > Microbial cell biology > Organelle isolation
Cell Biology > Organelle isolation > Extracellular vesicle
Biological Sciences > Biological techniques > Microbiology techniques
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.