- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

SarkoSpin: A Technique for Biochemical Isolation and Characterization of Pathological TDP-43 Aggregates

Published: Vol 9, Iss 22, Nov 20, 2019 DOI: 10.21769/BioProtoc.3424 Views: 7225
Reviewed by: Andrew L. Eagledeepika jaiswalEhsan KheradpezhouhAnonymous reviewer(s)
Original research article
The authors used this protocol in:
Jan 2019
Advertisement



Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Abstract
TDP-43 is the main aggregating protein in neurodegenerative diseases such as amyotrophic lateral sclerosis (ALS) and frontotemporal lobar degeneration (FTLD). Aggregated TDP-43 is resistant to diverse detergent solubilization, yet physiological TDP-43 and other abundant proteins commonly co-purify with pathological TDP-43. This mixed isolation has precluded the elucidation of the biochemical and structural features of the pathological TDP-43 and its role in disease. Here we describe SarkoSpin, a method for the isolation of pure pathological TDP-43 from patient autopsy brain by sample solubilization with Sarkosyl after nuclease treatment. This purification, which is also applicable to cell culture material, permits the study of biochemical properties of exclusively pathological TDP-43, allowing for the first time the determination of their link to the clinical presentation of FTLD. This method opens up a path for the study of pathological TDP-43 at the molecular and structural level in the heterogeneous spectrum of ALS and FTLD cases.
Keywords: TDP-43Background
Protein aggregation in affected neurons is a hallmark of neurodegenerative diseases (Soto et al., 2003). In two of the most devastating forms, amyotrophic lateral sclerosis (ALS) and frontotemporal lobar degeneration (FTLD), the principal aggregating protein is TAR-DNA binding protein 43 (TDP-43) (Ling et al., 2013). Under pathological conditions, TDP-43 is abnormally ubiquitinated, hyperphosphorylated and proteolytically cleaved, and is sequestered in insoluble cytoplasmic inclusions that include both the full-length protein as well as its highly-aggregation prone C-terminal fragment (Neumann et al., 2006; Arai et al., 2006). TDP-43 inclusions account for virtually all ALS patients and for up to 45% of FTLD patients, suggesting that ALS and FTLD are the two ends of a single clinical, genetic and pathological spectrum (Ling et al., 2013). FTLD cases can be subdivided into at least three subtypes (A, B and C) based on the shape and location of the aggregated TDP-43 (Lee et al., 2017), and it was recently shown that different biochemical features of the pathological TDP-43 may underlie this heterogeneity (Laferriere et al., 2019).
An RNA-binding protein (Buratti et al., 2001), TDP-43 forms physiological oligomers that are required to perform its indispensable role in RNA splicing regulation (Afroz et al., 2017). Pathological TDP-43 becomes resistant to solubilization by a wide range of detergents, but its isolation from physiological TDP-43 in brain tissue is hindered by protein-protein and protein-RNA interactions in the lysate. TDP-43 and other RNA-binding proteins containing low complexity domains may form homo- and heterooligomeric species in complex with nucleic acids that become partially insoluble.
Here, we describe SarkoSpin, a purification assay for the isolation of pathological TDP-43 from patient brain tissue by utilizing a single-step solubilization with nuclease treatment (Laferriere et al., 2019). This technique allows for the isolation of pure pathological TDP-43 from 99.8% of the total protein mass, including proteins that form big complexes in their physiological state, such as actin or histones. Isolated TDP-43 is pathologically modified and becomes polyubiquitination, hyperphosphorylated and C-terminally cleaved. Importantly, purified TDP-43 is free from the presence of other RNA-binding proteins containing low-complexity domains, such as hnRNPA1 (heterogeneous nuclear ribonucleoprotein A1), hnRNPH (heterogeneous nuclear ribonucleoprotein H), FUS (fused in sarcoma) or SOD1 (superoxide dismutase 1), which are also associated with ALS and FTLD. The isolation of pure fractions of pathological TDP-43 in combination with other high-resolution techniques will represent a major advancement in the understanding of the molecular mechanisms and structural differences of TDP-43 underlying ALS and FTLD heterogeneity.
Materials and Reagents
- Open-top tubes for an SW60 rotor. e.g., Open-top polyclear tubes 7/16x2-3/8" (Seton, catalog number: 7010)
- Filtered pipet tips (any commercially available are suitable, examples in brackets)
- 10 µl filter tip (STARLAB, catalog number: S1121-3810)
- 20 µl filter tip (STARLAB, catalog number: S1120-1810)
- 200 µl filter tip (STARLAB, catalog number: S1120-8810)
- 1000 µl filter tip (STARLAB, catalog number: S1122-1830)
- 15 ml tubes (any commercially available are suitable, e.g., Greiner Bio-One, catalog number: 188271)
- 50 ml tubes (any commercially available are suitable, e.g., Greiner Bio-One, catalog number: 227261)
- 6-well tissue culture plates (any commercially available are suitable, e.g., TPP catalog number: 92006)
- Protein LoBind tubes 1.5 ml (Eppendorf, catalog number: 0030108116)
- Divisible 250 µl 96-well PCR plates (Eppendorf, catalog number: 0030133374)
- N-Lauroylsarcosine sodium salt (Sarkosyl, Sigma, catalog number: L5125-100G)
- Trizma® hydrochloride (Tris-HCl, Sigma, catalog number: T3253-1KG)
- Sodium chloride (NaCl, Sigma, catalog number: 71380)
- EDTA disodium salt dihydrate (Amresco, catalog number: 0105-1KG)
- DL-Dithiothreitol (DTT, Thermo Scientific, catalog number: R0861)
- Phosphatase inhibitors. e.g., PhosSTOP (Roche, catalog number: 4906837001)
- Complete Mini EDTA-free Protease Inhibitor Cocktail (Roche, catalog number: 11836170001)
- Lysing kit for tissue homogenizing CKMix (Precellys, catalog number: P000918-LYSK0-A)
- Magnesium chloride (MgCl2, Sigma-Aldrich, catalog number: M8266-100G)
- Benzonase nuclease HC, purity > 90% 25KU (Merck, catalog number: 71205-3)
- Cell culture grade phosphate buffered saline (PBS Gibco, catalog number: 10010015)
- Sucrose (Millipore, catalog number: 84100)
- OptiPrepTM Density Gradient Medium (Sigma-Aldrich, catalog number: D1556)
- Dry ice
- Distilled water (Invitrogen, catalog number: 10977035)
- 2x Homogenization-solubilization (HS) buffer (see Recipes)
- 1x HS buffer (see Recipes)
- Benzonase mix (see Recipes)
- 2x HS buffer 4% Sarkosyl (w/v) (see Recipes)
- 1x HS buffer 0.5% Sarkosyl (w/v) (see Recipes)
- Benzonase mix for cell (see Recipes)
- Sucrose cushion (see Recipes)
- 8x HS buffer 4% Sarkosyl (w/v) (see Recipes)
- 2x HS buffer 1% Sarkosyl (w/v) (see Recipes)
Equipment
- Forceps (any commercially available are suitable, e.g., VWR, catalog number 232-1221)
- Scalpel (any commercially available is suitable, e.g., VWR, catalog number 233-5363)
- -80 °C freezer for storage of homogenate aliquots (any commercially available freezer is suitable)
- Minilys homogenizer (P000673-MLYS0-A, Bertin technologies)
- Bucket for dry ice (any commercially available is suitable, e.g., Fisher Scientific, catalog number, 11324085)
- Thermo Mixer C (Eppendorf, catalog number: 5382000015) with SmartBlock 1.5 ml (Eppendorf, catalog number: 5360000038)
- Bench top centrifuge. e.g., PrismTM Microcentrifuge (Labnet International, C2500)
- Gradient Master Base Unit (BioComp, #108) with 7/16" (11 mm) MagnabaseTM holder and marker block for SW60 rotor short caps (BioComp, #105-911A-R)
- Ultracentrifuge. e.g., OPTIMATM XPN-100 ultracentrifuge (Beckman Coulter, A99846)
- SW 60 Ti Swinging-Bucket Rotor (Beckman Coulter, 335649)
- Vortex (any commercially available freezer is suitable; e.g., Scientific Industries, SI-0236)
- Micropipettes (any commercially available are suitable, examples in brackets)
P10 (Eppendorf, catalog number: 3120000020)
P20 (Eppendorf, catalog number: 3120000038)
P200 (Eppendorf, catalog number: 3120000054)
P1000 (Eppendorf, catalog number: 3120000062) - Piston gradient fractionator (BioComp, 152)
- Fraction collector (Gilson, FC203B)
Procedure
- Homogenization of tissue (Figure 1A)

Figure 1. Schematic representation of the SarkoSpin protocol. A. Brain autopsy tissue is mechanically homogenized and divided into single-use aliquots, which are snap-frozen for future uses. B. Aliquoted homogenates are solubilized in a sarkosyl-based buffer and treated with nuclease in order to disrupt major protein-protein and protein-RNA complexes, before C. the bulk of insoluble pathological TDP-43 can be isolated in a single spin. (D, E) Alternatively, the biochemical properties of the insoluble pathological TDP-43 can be analyzed by different analytical centrifugation methods.- Take the patient samples out of the -80 °C freezer into a box containing dry ice to avoid thawing of the tissue while processing.
- Place the brain sample on the balance over a paper and let it thaw slightly.
- Cut a piece with a maximum weight of 300 mg from the brain sample with a sterile scalpel and autoclaved forceps.
- Return the remainder of the sample into dry ice to avoid further thawing and damage of the tissue.
- Chop the brain sample on the balance into smaller portions with the sterile scalpel.
- For homogenization, transfer the tissue pieces into the 2 ml-tube from the lysing kit, containing a mixture of ceramic beads with a diameter of 1.4 and 2.8 mm.
- Add HS buffer in a ratio of 5:1 to the amount of tissue. For example, for the maximum of 300 mg per tube, add 1.5 ml 1x HS buffer (Recipe 2).
- Homogenize the samples in a Minilys device three times for 30 s each at maximum speed. Cool down the sample on ice between rounds.
- Let the sample set down for the froth to disappear. A short spin of the tube in a table-top microfuge will help.
- Vortex quickly at low speed while avoiding creating new froth.
- Divide the homogenate into aliquots of 150 µl in low protein binding 1.5 ml tubes.
- Keep 10 µl aliquot from the original homogenate to check levels of phosphorylated TDP-43 in the sample by Western blot and 5 µl for protein concentration determination (directly add to 45 µl 1x HS buffer to dilute it). This step will allow determining whether homogenates indeed have comparable starting protein concentrations, and therefore SarkoSpin the on homogenates from different brain samples will be comparable.
- Snap-freeze the 150 µl aliquots on dry ice and store at -80 °C. Homogenate aliquots are good for SarkoSpin purification for months to years if properly stored at -80 °C.
- Solubilization (Figure 1B)
- Thaw the homogenate aliquots (from Step A13) on ice.
- Add 50 µl of Benzonase mix (Recipe 3) to each sample.
- Incubate at room temperature for 5 min.
- Add 200 µl of 2x HS buffer 4% Sarkosyl (Recipe 4) to each sample.
- Incubate in a Thermo Mixer at 38 °C and shaking at 600 rpm for 45 min.
When performing SarkoSpin from cultured cell samples, cell lysis and solubilization are performed in a combined step as follows:- Per well of a 6-well plate, lyse cells in 100 µl 1x HS buffer 0.5% Sarkosyl.
- Add 5 µl of Benzonase mix for cells (Recipe 3).
- Incubate for 5 min at RT.
- Scrape the cells and transfer the lysate to a low protein binding 1.5 ml tube.
- Wash the well with additional 100 µl of 1x HS buffer 0.5% Sarkosyl and transfer to the tube.
- Vortex to homogenize.
- Transfer 170 µl of the lysate into a new low protein binding 1.5 ml tube, leaving 5 µl for protein concentration determination and 30 µl for Western blot analysis of total lysate.
- Add 178 µl 2x HS 4% Sarkosyl and 1x HS to 170 µl of lysate, for a final concentration of 2% Sarkosyl in a volume of 400 µl (like for brain homogenates solubilization).
- Incubate on ice for 45 min, vortexing in intervals of 10 min.
- Proceed with the rest of the protocol as for the brain homogenates.
- Isolation (Figure 1C)
- Add 200 µl of ice-cold 1x HS buffer 0.5% Sarkosyl (Recipe 5) to each sample.
- Centrifuge at 21,200 x g for 30 min at room temperature (RT) in a bench top centrifuge.
- Label low protein binding 1.5 ml tubes with the sample name and “Supernatant/“Pellet”.
- After the centrifugation, remove up to 500 µl of the supernatant and keep it aside in the tube pre-labeled as “Supernatant”.
- Carefully remove the remaining supernatant with a gel-loading tip and discard.
- Let the sample stand for one minute and repeat Step C5 to ensure that all supernatant is removed.
- Carefully wash the pellet (brown) and tube walls with 100 µl cell culture grade PBS. Lipids can appear as a white layer above the protein pellet and can be carefully washed away with the PBS (for complete removal of lipids in the sample, see below).
- Remove all traces of PBS by repeating Steps C5 and C6.
- Resuspend the pellet in the desired amount of 1x HS buffer 0.5% Sarkosyl and transfer to the tube pre-labeled as “Pellet”.
If samples are not going to be immediately used for analysis, do not resuspend in the pellet. Snap-freeze the samples on dry ice and store at -80 °C. - Proceed to pellet analysis by SDS-PAGE, native PAGE or dot blot and Western blot (Data analysis, Figure 2).
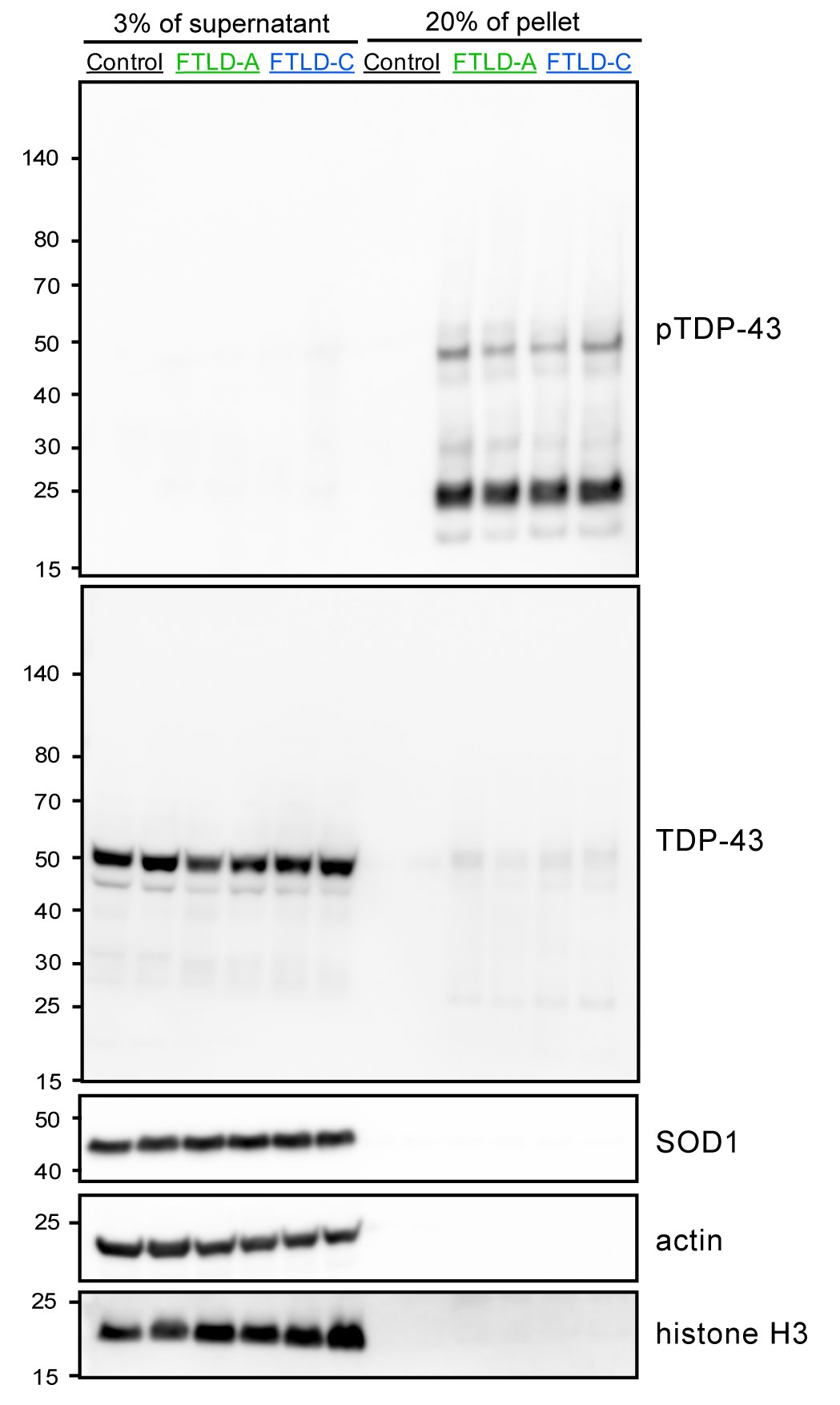
Figure 2. Representative Western blots of the supernatant and pellet fractions from SarkoSpin isolation. Pathological phosphorylated TDP-43 is enriched in the pellet fraction of samples from FTLD patients but not in healthy controls, while the bulk of the physiological TDP-43 remains soluble. Importantly, abundant cellular proteins such as SOD1, actin and histone H3 also remain in the supernatant fraction.
- Lay 400 µl of solubilized homogenates over 400 µl of sucrose cushion (Recipe 7) in low protein binding 1.5 ml tubes.
- Centrifuge at 21,200 x g for 30 min at room temperature (RT) in a bench top centrifuge.
- Proceed with the protocol as described from Step C3 on.
- Velocity sedimentation assay (Figure 1D, continues from Step B5)
- Prepare the OptiPrep gradient cushions as indicated in Table 1.
Table 1. Recipe for the OptiPrep gradient cushions required for the velocity sedimentation assay. For 2x HS buffer 1% Sarkosyl, see Recipe 9. For the commercial OptiPrep solution (60%), see Materials and Reagents.
- Use a Gradient Master to prepare the linear OptiPrep gradient in 4.2 ml SW60 rotor tubes as follows:
- With the tube in the marker block for an SW 60 Ti rotor, draw a line at the middle of the tube for short caps.
- Pipette the 10% OptiPrep cushion into the tube until a few mm above the drawn line.
- Carefully inject the 25% OptiPrep cushion at the bottom of the tube with the provided steel cannula, thereby pushing the previous 10% OptiPrep cushion up, until the interface reaches the mid-tube mark.
- Close the tube with the provided short cap, removing the remaining air inside.
- Use the scroll and spirit level on the Gradient Master in order to make the Magnabase horizontal.
- Place the tubes in the Magnabase, and transfer onto the Gradient Master plate.
- Run the program: SW60-1 step://1 min 10 s//82º //20 rpm//.
- Once finished, carefully remove the tube caps and remove approximately 150 µl of the upper layer of the gradient.
- Weigh the tubes containing the gradients and balance them as precisely as possible (to the decigram level).
- Carefully lay 200 µl of the solubilized homogenate (Step B5) over the top of the gradient.
- Ultracentrifuge at 400,000 x g for 45 min at 4 °C with maximum acceleration and deceleration.
- Carefully remove the tubes from the centrifuge buckets with tweezers and keep them at 4 °C before fractionation with a piston fractionator associated with a fraction collector.
- Insert a divisible 250 µl PCR in the collector holder.
- Rinse the piston before and after each gradient by fractionating of a tube filled with water and using the “rinse” button.
- Air dry the piston after the washing steps by opening the air screw and pressing the button Air to make it blow through the piston. Screw it back gently after drying.
- Place the gradient to fractionate in the tube holder, clipped with the cap.
- Let the piston go down with the position roll until the liquid reaches the collector head.
- Reset position and enter the program the following program: Speed: 0.5/Distance: 2.96/Number: 16
- Press Start. The fractionator will automatically fractionate the gradient into 16 fractions in the divisible 96-well PCR plate.
- Repeat the washing and drying (Steps D7 and D8) before fractionating another gradient.
- Prepare the OptiPrep gradient cushions as indicated in Table 1.
- Density flotation assay (Figure 1E, continues from Step B5)
- Prepare the OptiPrep gradient cushions as indicated in Table 2.
Table 2. Recipe for the OptiPrep gradient cushions required for the density flotation assay. For 8x HS buffer 4% Sarkosyl, see Recipe 8. For the OptiPrep solution (60%), see Materials and Reagents.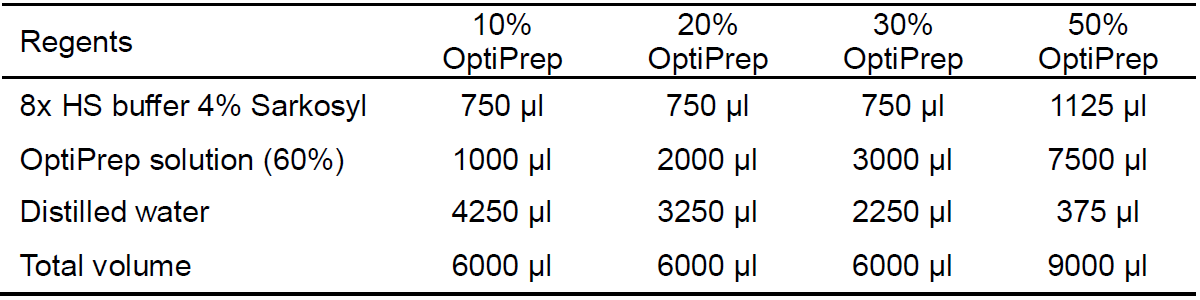
- In order to prepare the discontinuous OptiPrep gradient, carefully lay the OptiPrep cushions in 4.2 ml SW60 rotor tubes to create the discontinuous gradient in the following order (highest concentration at the bottom):
300 μl of pure (60%) OptiPrep solution
700 μl of the 50% OptiPrep cushion
800 μl of the mix (220 μl of solubilized homogenate from step B5 + 680 μl 50% OptiPrep cushion) for a final OptiPrep concentration of 40%
800 μl of the 30% OptiPrep cushion
800 μl of the 20% OptiPrep cushion
800 μl of the 10% OptiPrep cushion
Weigh the tubes after pipetting each cushion to balance them to the decigram level. - Ultracentrifuge at 125,000 x g for 17 h at 4 °C with maximum acceleration and deceleration.
- Fractionate as describe from Step D5.
- Prepare the OptiPrep gradient cushions as indicated in Table 2.
Data analysis
Depending on the application of the SarkoSpin isolation, insoluble TDP-43 can be analyzed with a wide variety of techniques, ranging from Western blots (after SDS-PAGE, native PAGE or filter retardation assay, Figure 2) to high-resolution techniques like electron microscopy or mass spectrometry (Laferriere et al., 2019).
Recipes
- 2x Homogenization-solubilization (HS) buffer
20 mM Tris, pH 7.5
300 mM NaCl
1 mM EDTA
2 mM DTT, with protease and phosphatase inhibitors
Stable at 4 °C for one month without inhibitors
For 50 ml:
1 ml of 1 M Tris-HCl, pH 7.4 (78.8 g Tris-HCl in 500 ml H2O, filtered-sterilized and stored at RT)
6 ml of 2.5 M NaCl (36.53 g NaCl in 250 ml H2O, filtered-sterilized and stored at RT)
100 µl of 0.5 M EDTA, pH 8 (93.06 g EDTA disodium salt dihydrate in 500 ml H2O, pH adjusted to 8, filtered-sterilized and stored at RT)
100 µl of 1 M DTT (1.5 g of DTT in 8 ml of H2O, aliquoted in single-use amounts and stored at -20 °C)
Fill up with distilled water to 50 ml (approx. 42.8 ml)
For every 10 ml of 2x HS buffer, 2 tablets Complete Mini EDTA-free and 1 tablet PhosSTOP - 1x HS buffer
10 mM Tris, pH 7.4, 15 mM NaCl, 0.5 mM EDTA, 1 mM DTT, with protease and phosphatase inhibitors
For 10 ml:
5 ml 2x HS buffer
5 ml distilled water - Benzonase mix
12 mM MgCl2 1x HS buffer with 250 U/sample Benzonase
For 50 µl:
1 µl Benzonase HC
50 µl 1x HS buffer
1.4 µl 500 mM MgCl2 (2.38 g MgCl2 in 50 ml H2O, filtered-sterilized and stored at 4 °C) - 2x HS buffer 4% Sarkosyl (w/v)
400 mg Sarkosyl per 10 ml of 2x HS buffer - 1x HS buffer 0.5% Sarkosyl (w/v)
50 mg Sarkosyl per 10 ml of 1x HS buffer - Benzonase mix for cells
60 mM MgCl2 1x HS buffer with 100 U/well Benzonase
For 6 µl:
0.4 µl Benzonase HC
5 µl 1x HS buffer
0.6 µl 500 mM MgCl2 - Sucrose cushion
40% sucrose (w/v) in 1x HS buffer
For 400 µl:
160 mg sucrose
Fill up to 400 µl with 1X HS buffer - 8x HS buffer 4% Sarkosyl (w/v)
80 mM Tris, pH 7.5, 1.2 M NaCl, 4 mM EDTA, 8 mM DTT, with protease inhibitors and 4% Sarkosyl (w/v). Stable at 4 °C for one month without inhibitors (f)
For 4 ml:
320 µl of 1 M Tris-HCl pH 7.4
960 µl of 2.5 M NaCl
32 µl of 0.5 M EDTA pH 8
32 µl of 1 M DTT
Fill up with distilled water to 4 ml (approx. 2656 µl)
1 tablet cOmplete Mini EDTA-free
160 mg Sarkosyl - 2x HS buffer 1% Sarkosyl (w/v)
100 mg Sarkosyl per 10 ml of 2x HS buffer
Acknowledgments
M. P. is the recipient of a Swiss National Science Foundation Professorship (PP00P3_144862), a Human Frontier Science Program Career Development Award (CDA-00058/2012) and a UCL/ZNZ Neuroscience Collaboration Grant funded through UCL’s Welcome Trust Institutional Strategic Support Fund Investing in Excellent Researchers (105604/Z/14/Z). M.P.-B. received a Candoc Award (Forschungskredit) from the University of Zurich. F.L. is supported by the Milton-Safenowitz fellowship from the ALS Association (15-IIP-208) and a Postdoc Award from the University of Zurich.
This protocol is based on the publication Laferriere et al., 2019.
Competing interests
The authors declare no competing financial interests.
Ethics
The Queen Square Brain Bank for Neurological disorders (UCL Institute of Neurology), the Department of Neurology (University of California San Diego) or the Netherlands Brain Bank (Netherlands Institute for Neuroscience) provided the brain autopsy material. Brain tissue was donated upon written informed consent from all subjects and the respective institution obtained the clinical information for research purposes.
References
- Afroz, T., Hock, E. M., Ernst, P., Foglieni, C., Jambeau, M., Gilhespy, L. A. B., Laferriere, F., Maniecka, Z., Pluckthun, A., Mittl, P., Paganetti, P., Allain, F. H. T. and Polymenidou, M. (2017). Functional and dynamic polymerization of the ALS-linked protein TDP-43 antagonizes its pathologic aggregation. Nat Commun 8(1): 45.
- Arai, T., Hasegawa, M., Akiyama, H., Ikeda, K., Nonaka, T., Mori, H., Mann, D., Tsuchiya, K., Yoshida, M., Hashizume, Y. and Oda, T. (2006). TDP-43 is a component of ubiquitin-positive tau-negative inclusions in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Biochem Biophys Res Commun 351(3): 602-611.
- Buratti, E., Dork, T., Zuccato, E., Pagani, F., Romano, M. and Baralle, F. E. (2001). Nuclear factor TDP-43 and SR proteins promote in vitro and in vivo CFTR exon 9 skipping. EMBO J 20(7): 1774-1784.
- Laferriere, F., Maniecka, Z., Perez-Berlanga, M., Hruska-Plochan, M., Gilhespy, L., Hock, E. M., Wagner, U., Afroz, T., Boersema, P. J., Barmettler, G., Foti, S. C., Asi, Y. T., Isaacs, A. M., Al-Amoudi, A., Lewis, A., Stahlberg, H., Ravits, J., De Giorgi, F., Ichas, F., Bezard, E., Picotti, P., Lashley, T. and Polymenidou, M. (2019). TDP-43 extracted from frontotemporal lobar degeneration subject brains displays distinct aggregate assemblies and neurotoxic effects reflecting disease progression rates. Nat Neurosci 22(1): 65-77.
- Lee, E. B., Porta, S., Michael Baer, G., Xu, Y., Suh, E., Kwong, L. K., Elman, L., Grossman, M., Lee, V. M., Irwin, D. J., Van Deerlin, V. M. and Trojanowski, J. Q. (2017). Expansion of the classification of FTLD-TDP: distinct pathology associated with rapidly progressive frontotemporal degeneration. Acta Neuropathol 134(1): 65-78.
- Ling, S. C., Polymenidou, M. and Cleveland, D. W. (2013). Converging mechanisms in ALS and FTD: disrupted RNA and protein homeostasis. Neuron 79(3): 416-438.
- Neumann, M., Sampathu, D. M., Kwong, L. K., Truax, A. C., Micsenyi, M. C., Chou, T. T., Bruce, J., Schuck, T., Grossman, M., Clark, C. M., McCluskey, L. F., Miller, B. L., Masliah, E., Mackenzie, I. R., Feldman, H., Feiden, W., Kretzschmar, H. A., Trojanowski, J. Q. and Lee, V. M. (2006). Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science 314(5796): 130-133.
- Soto, C. (2003). Unfolding the role of protein misfolding in neurodegenerative diseases. Nat Rev Neurosci 4(1): 49-60.
Article Information
Copyright
© 2019 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Pérez-Berlanga, M., Laferrière, F. and Polymenidou, M. (2019). SarkoSpin: A Technique for Biochemical Isolation and Characterization of Pathological TDP-43 Aggregates. Bio-protocol 9(22): e3424. DOI: 10.21769/BioProtoc.3424.
Category
Neuroscience > Nervous system disorders > Neurodegeneration
Biochemistry > Protein > Isolation and purification
Molecular Biology > Protein > Detection
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.