- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Measuring Spatiotemporal Dynamics of Odor Gradient for Small Animals by Gas Chromatography

Published: Vol 8, Iss 7, Apr 5, 2018 DOI: 10.21769/BioProtoc.2797 Views: 7537
Reviewed by: Neelanjan BoseSanjib GuhaKarthik Krishnamurthy
Original research article
The authors used this protocol in:

May 2017
Advertisement



Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics








Abstract
Odor is the most fundamental chemical stimulus that delivers information regarding food, mating partners, enemies, and danger in the surrounding environment. Research on odor response in animals is widespread, although studies on experimental systems in which the gradient of odor concentration is quantitatively measured has been quite limited. Here, we describe a method for measuring a gradient of odor concentration established by volatilization and diffusion in a relatively small enclosed space, which has been used widely in laboratories to analyze small model animals such as the nematode Caenorhabditis elegans and the fruit fly Drosophila melanogaster. We first vaporized known amounts of a liquid odorant 2-nonanone in a tank and subjected them to gas chromatographic analysis to obtain a calibration curve. Then, we aspirated a small amount of gas phase from a small hole on an agar plate and measured the odor concentration. By repeating this at different spatial and temporal points, we were able to detect a gradient of the odor concentration that increased over time. Furthermore, by applying these measured values to mathematical models of volatilization and diffusion, we were able to visualize an estimated dynamic change in odor concentration over an agar plate. Combining monitoring of odor concentration change in an agar plate with behavioral monitoring by machine vision will allow us to estimate how the brain computes information regarding odor concentration change in order to regulate behavior.
Keywords: OdorantBackground
Odor is the most fundamental chemical stimulus that conveys the existence of food, reproductive partners, enemies, etc. in the surrounding environment. Small model animals, such as the nematode Caenorhabditis elegans and fruit fly Drosophila melanogaster are suitable for understanding brain responses to odor stimuli at the levels of behavior, neural activity, and molecules because: (1) behavioral responses to odor stimuli can be easily recorded with inexpensive high resolution cameras; (2) responses in multiple neurons/neuronal groups can be measured with calcium imaging and (3) genes responsible for behavioral and neural responses can be identified with various genetic methods (De Bono and Maricq, 2005; Venken et al., 2011).
However, it is difficult to measure the odor concentrations that are actually sensed by these small animals during their behavior in a small arena suited for observation. In general, the measurement of odor concentration requires constant air flow in a device supplying the odorant-containing air to the sensor. Thus, air should be constantly drawn from the air phase of the arena, destroying the odor gradient. Measuring odor gradient in a small behavioral arena has been achieved either by strengthening the air flow for the odor gradient compared to the flow for sampling or by optically measuring the air phase odorant concentration. Gershow et al. (2012) developed a relatively large apparatus (30 x 30 cm) for Drosophila larvae for slow but large (2 L/min) constant parallel flows with different concentrations, in order to create an odor concentration gradient perpendicular to the flow. Louis et al. (2008) used infrared beams to measure integrated concentrations of odor on one axis in a naturally evaporated and diffused gradient, and calculated the gradient shape mathematically based on Gaussian diffusion. The former method allows quantitative measurement of the odor gradient, although it is not based on natural evaporation and diffusion. It also requires specific, controlled apparatus. The latter method is suitable for natural gradients, although it does not allow accurate measurement of specific positions.
Here we report a method to measure a dynamic odor gradient in a widely-used plastic plate by gas chromatography (GC). Observing odor-taxis behaviors on plastic plates with an agar layer is easy and thus is conducted in many laboratories. In addition, we are able to video-record the behaviors using inexpensive USB cameras. Therefore, by measuring temporal changes in the odor gradient on an agar plate, we can obtain clues to estimate brain computations controlling how temporal changes in odor stimuli affect the animal’s behavior.
For measurement, first, specific amounts of liquid odorant are individually volatilized in a vaporizing tank to make gas with known concentrations of odorant. Then, the gas is subjected to GC with different concentrations, in order to calculate a calibration curve for known gas concentrations and GC values. Next, a small amount of gas is sampled from a specific spatio-temporal point on an agar plate with evaporating and diffusing odorant, and subjected to GC analysis. Finally, the entire odor gradient is calculated by the measured concentrations at different spatio-temporal points. In our experiment, measurements suggested that C. elegans responds behaviorally to odor concentration changes as small as ± 0.01 μM/sec in ~2 μM concentration on a natural odor gradient. This is consistent with results from an experiment with artificial and controlled odor concentration changes (Tanimoto et al., 2017).
Materials and Reagents
- ø 9 cm sterile Petri dish (IWAKI, catalog number: SH90-15 ) with nematode growth medium (NGM) agar
Note: Pour 10 ml of autoclaved 1.5-2.5% agar solution per dish following the regular sterilized technique to make an agar plate for behavioral analysis. We used NGM agar for C. elegans behavioral analysis. This agar plate can be stored at 4 °C for a few weeks. The plates should be moved to a bench a few hours before the assay and kept without their lids for 15-30 min to dry. Dried plates with lids are placed upside-down on a bench. The plates are not sealed with either Parafilm or sticky tape. - Microliter syringe, 50 μl, cemented needle (Hamilton, catalog number: 80565 )
Note: This is a blunt needle point. - Micro-volume syringe, 5 μl, fixed needle (SGE, catalog number: 001000 )
- Plastic disposable syringe, 2.0 ml (Top, catalog number: 5079-01 )
- Replacement needle (Luer lock side hole), 23 G x 4 cm (GL Sciences, catalog number: 3008-46004 )
- Pasteur pipette (IWAKI, catalog number: 1K-PAS-5P )
- Dropper bulb (AS ONE, catalog number: 1-6227-05 )
- 2-Nonanone (Wako Pure Chemical Industries, catalog number: 132-04173 )
Note: Liquid at room temperature. Although we used only this odorant, this protocol could be used for other odorants as well. - EtOH (Wako Pure Chemical Industries, catalog number: 057-00456 )
- Sodium chloride (NaCl) (Wako Pure Chemical Industries, catalog number: 191-01665 )
- Bacto peptone (BD, BactoTM, catalog number: 211677 )
- Agar (Wako Pure Chemical Industries, catalog number: 010-08725 )
- Cholesterol (Wako Pure Chemical Industries, catalog number: 034-03002 )
- Calcium chloride dihydrate (CaCl2·2H2O) (Wako Pure Chemical Industries, catalog number: 038-12775 )
- Magnesium sulfate heptahydrate (MgSO4·7H2O) (Wako Pure Chemical Industries, catalog number: 131-00405 )
- Dipotassium hydrogenphosphate (K2HPO4) (Wako Pure Chemical Industries, catalog number: 164-04295 )
- Potassium dihydrogen phosphate (KH2PO4) (Wako Pure Chemical Industries, catalog number: 169-04245 )
Equipment
- Vaporizing tank (FIS, catalog number: DT-T1 ) (Figure 1)
Note: A custom-made acrylic tank of 50 L, equipped with a small metal block with a vaporizing groove, a heater with a temperature controller, and a fan. The odorant liquid is placed in the groove of the metal block through a liquid inlet on the lid, and the metal block is warmed with the heater to facilitate volatilization of the odorant. The fan stirs the air so that the volatilized odorant is distributed equally in the tank.
Figure 1. Vaporizing tank - Gas chromatograph (GC) (Nissha FIS, model: SGVA-N2 ) (Figure 2)
Note: A simple and inexpensive GC optimized for 2-nonanone with a semiconductor detector. Other GC can also be used.
Figure 2. Gas chromatograph - Carrier gas cylinder (Air Liquide, model: Alphagaz 1 )
Note: This was recommended by Nissha FIS Inc. - Vacuum cleaner (Toshiba, catalog number: VC-PC6A , L)
Note: A domestic vacuum cleaner. - Pin vise (Tamiya, Fine Pin Vise D [0.1-3.2 mm])
- 1 mm drill (Tamiya, Basic Drill Bit set 5 pc)
Software
- SGC.exe (Nissha FIS Inc., Hyogo, Japan)
Note: This is a specific software to control this type of GC provided by the manufacturer. Software should be installed on a Windows PC (XP, Vista, or 7) that is connected to the GC. Depending on the GC instrument being used, the manufacturer may have specific analysis software recommendations.
Procedure
Overview: We vaporized specific amounts of liquid 2-nonanone in the vaporizing tank to make 2-nonanone gas of known concentrations, and measured GC values to calculate a calibration curve. Next, a gas phase of 0.2 ml in an agar plate was sampled and measured with the GC. Although we did the following with 2-nonanone, our method should be applicable for other odorants that are vaporized from the liquid at room temperature and can be measured by gas chromatography. Important steps are summarized in Video 1.

- Measuring known concentrations of 2-nonanone gas for a calibration curve
2-nonanone gas of known concentration can be obtained by vaporizing a specific amount of 2-nonanone liquid in the vaporizing tank (see below). A calibration curve can be obtained by measuring different concentrations of the gas with GC and correlating the measured values. However, the time to reach maximum odor concentration varied for each amount of liquid, likely due to differences in vaporization, diffusion, and trace adhesion to the vaporizing tank wall. Therefore, we monitored temporal changes in odor concentration in the tank to find the optimal time for vaporization of each concentration.- Flow the carrier gas at 0.25-0.35 MPa from the air cylinder connected to the GC.
Note: Do this immediately before turning on the GC. - Turn on the GC immediately after Step A1.
Note: Flowing gas without turning on power will damage the column inside the GC. - Wait until the ‘Ready’ lamp is illuminated.
Notes:- This may take about 90 min.
- If the GC is used after a long interval (e.g., more than 2 weeks), the measured value tends to be higher. In that case, use the GC a few days before taking actual measurements. An interval of up to several days has no effect.
- The high values recorded after long intervals are attributed to the following: During the interval, the sensor surface is coated with various small compounds in the air. Electrical conduction increases the temperature of the sensor to 300-400 °C, which clarifies the attached compounds and causes transient high sensitivity for several hours.
- The directions above are specific to the GC instrument used in this study. Other instrumentation may require adapted methods and steps.
- This may take about 90 min.
- Turn on the electrical switch of the vaporizing tank (Figure 1), and set the temperature at 50 °C: It will take about 10 min to reach 50 °C.
- Take an appropriate amount of 2-nonanone liquid (Table 1) with a glass syringe of 5 μl or 50 μl, insert it in the liquid inlet on the lid of the tank, and place the liquid in the vaporizing groove installed on the back side of the lid.
Table 1. Volumes of 2-nonanone liquid for the vaporization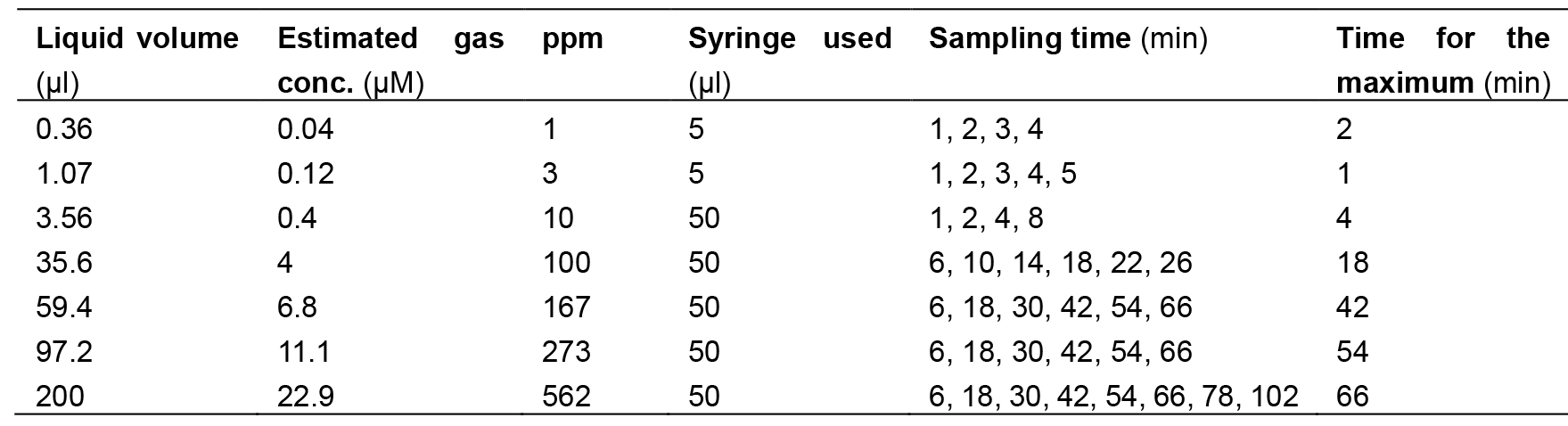
- The relationship between the liquid volume and the estimated gas concentration in Table 1 is as follows:

where C is the required gas concentration (mol/L), Vliquid is the amount of liquid (ml) to be added, d is the density of liquid odorant (g/ml), M is the molecular weight (g), and Vtank is the volume of the vaporizing tank (L). - Start SGC.exe on a Windows PC connected to the GC and operate according to the manual.
- Press the start button in SGC.exe.
- After a certain period of time (see Table 1), carefully insert the replacement needle attached to the disposable syringe from the gas outlet. Extract 0.2 ml and quickly remove the needle from the tank. Carefully insert the needle in the gas inlet of the GC (Figure 2) until it hits the bottom, and immediately infuse the gas inside the syringe.
Note: This step should be completed in about 5-6 sec. - Measurement is started by gas injection. Drawing of the graph (Figure 3) starts and ends automatically. Data is automatically saved. In the default setting, a measurement takes 8 min. The file can be exported as a CSV file.
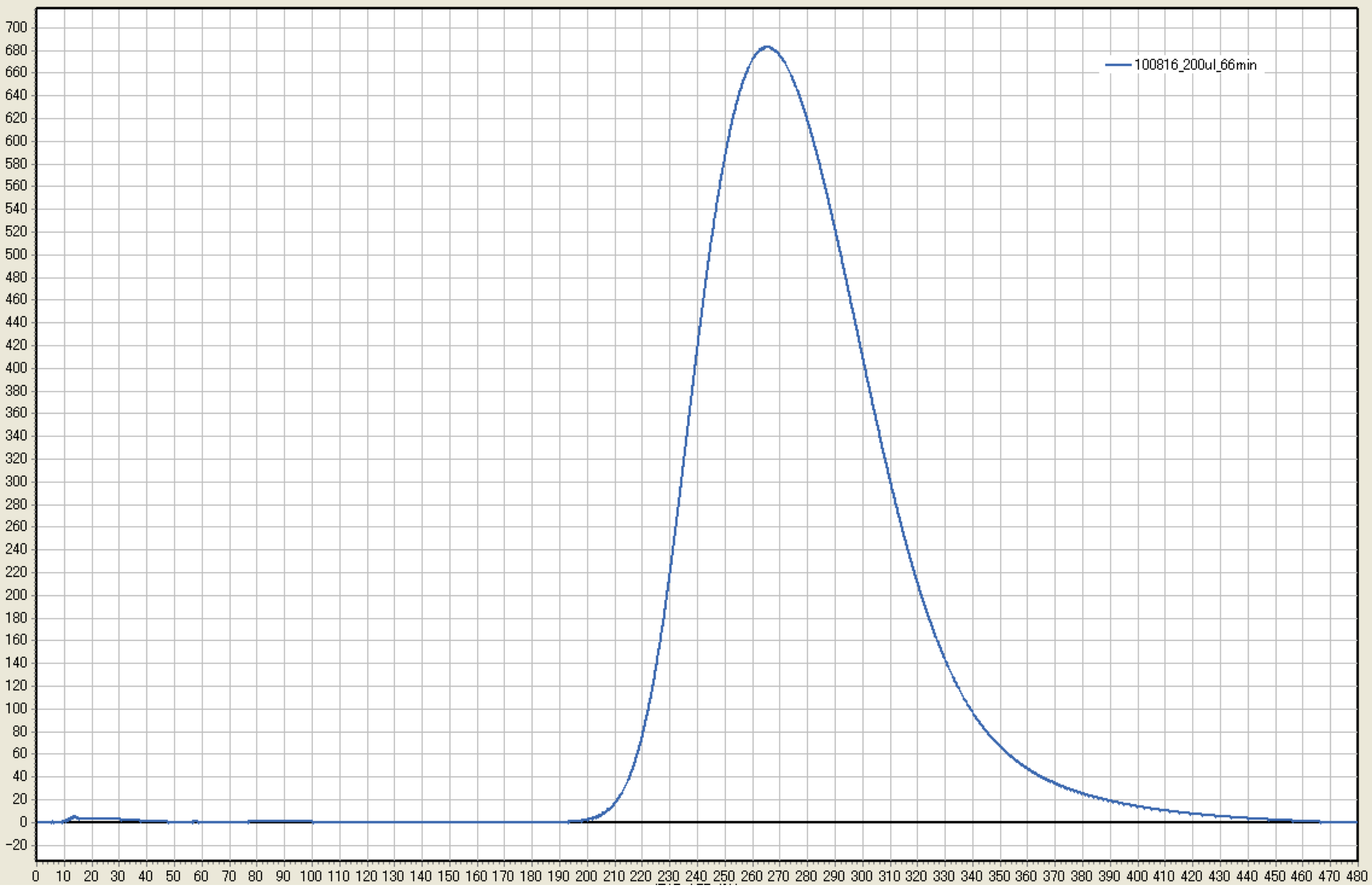
Figure 3. A representative result of one measurement for 22.9 μM (i.e., 200 μl) of 2-nonanone. The horizontal axis is time (sec), and the vertical axis is the signal (mV). - To measure temporal changes in the measured value for the specific amount of the odorant, odor gas sampling can be performed at different time points for one odorant injection (e.g., 6, 18, 30, and 42 min for 6.8 and 11.1 μM; see Table 1). However, if there is no 8-min interval (e.g., 1, 2, 3, and 4 min for 0.04 and 0.12 μM), clean the tank (see the next section) and start from the gas vaporization.
- If gas remains in the tank (likely by adhesion to the wall), vaporization of the residual amount affects the GC value, especially when a small concentration is being measured. In order to avoid this, gas in the tank is removed with a vacuum cleaner, and the wall is wiped with a paper towel containing EtOH, followed by further suction with the vacuum cleaner. After this procedure, remaining gas was not detected in our experiment.
- For all conditions, repeat the measurement 3-4 times (once daily and repeat over 3-4 days) and find the time at which the average value is at a maximum (Figure 4). In the case of 2-nonanone, the relationship between odor concentrations and measured values are nicely fitted to two regression lines for concentrations lower and higher than 4 μM (R2 = 0.9991 and 0.9995, respectively; Figure 5) (Tanimoto et al., 2017). In general, for semiconductor detectors, the correlation between the peak height of the signal and signal concentration in a log-log plot is well-fitted by two simple regression lines for lower and higher concentrations. Therefore, these results are adopted as calibration curves for low and high concentrations. Excel (Microsoft) is used for data analysis.
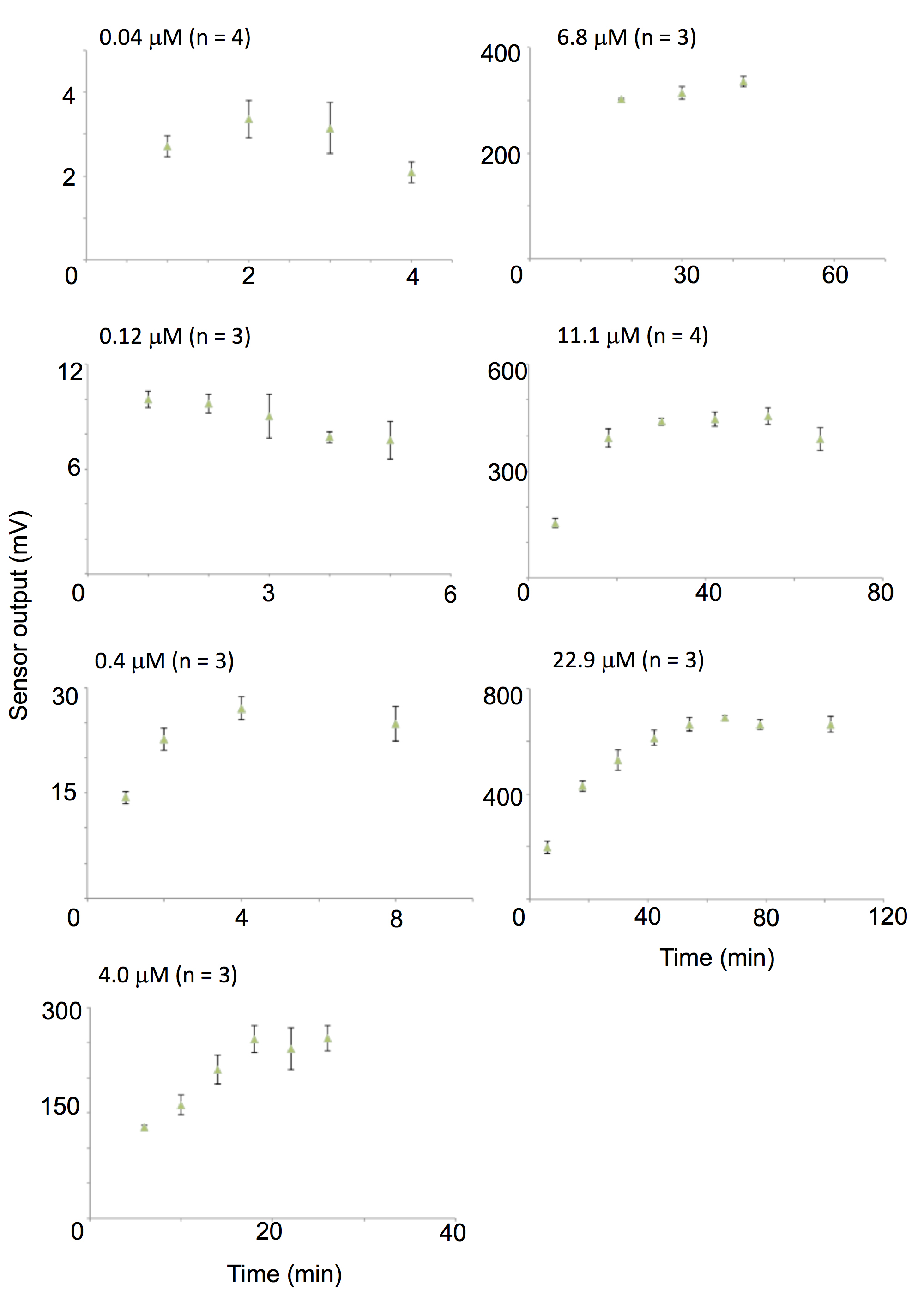
Figure 4. Changes over time of the measured value for different amounts of liquid odor. On each graph, the expected 2-nonanone gas concentration at saturation is shown. The horizontal (vaporizing time) and vertical (sensor output) axes are different in each graph. For 2-nonanone gas with a low saturation concentration of 0.04-0.12 μM, the concentration became saturated immediately and then decreased slightly. This is likely because of adhesion of 2-nonanone to the wall. For each condition, results are shown as mean ± standard error of 3-4 repetitions.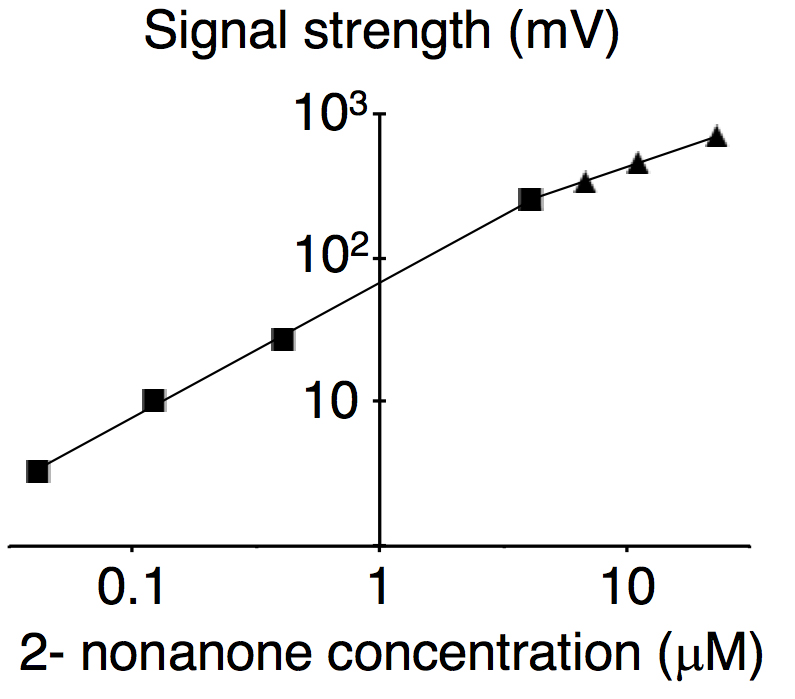
Figure 5. Calibration curve for 2-nonanone. Each dot represents average values of 3-4 experiments, and data on the log-log plot were fitted with two simple regression lines for lower (squares) and higher (triangles) concentrations. This figure was originally published in Tanimoto et al. (2017).
- Flow the carrier gas at 0.25-0.35 MPa from the air cylinder connected to the GC.
- Measuring 2-nonanone gradient on an agar plate
- On the back side of the agar plate, mark the positions of odor spot and gas sampling with a pen.
Note: We measured at six points (x, y, z) on the assay plate shown in Figure 6 at 1, 3, 6, 9, and 12 min. The motivations for adopting 6 points (x, y) for measurement are as follows:- Since we examined odor avoidance behavior of C. elegans, the four points on the x axis were chosen to measure the direction avoided by C. elegans, i.e., to measure the spatial gradient along the x direction. In the range of x > 0 in which C. elegans mainly existed, three points were chosen. Only one point was measured in the range x < 0 where C. elegans did not often exist.
- To measure the spatial gradient in the y direction, (22, 15) was selected. We chose only one point because the worms did not spread much along the y axis.
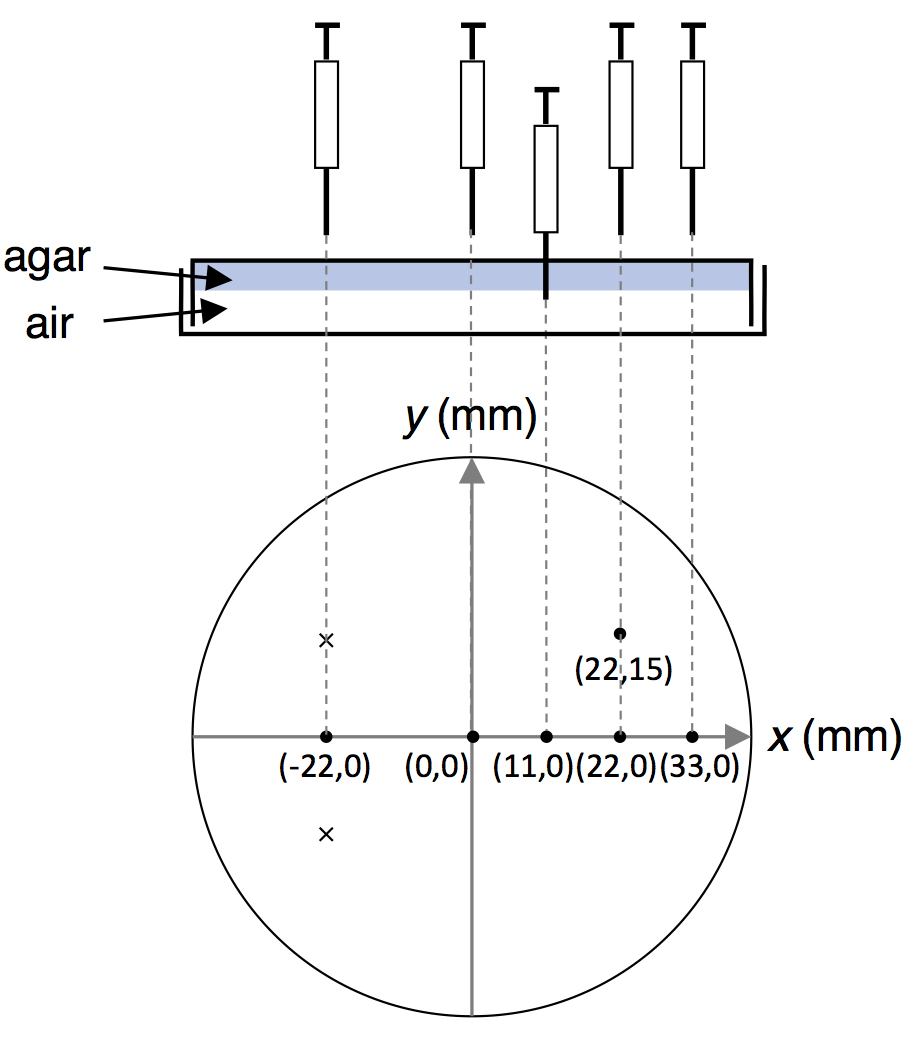
Figure 6. Gas sampling from the agar plate [originally published in Tanimoto et al. (2017). Creative Commons Attribution License]
- Since we examined odor avoidance behavior of C. elegans, the four points on the x axis were chosen to measure the direction avoided by C. elegans, i.e., to measure the spatial gradient along the x direction. In the range of x > 0 in which C. elegans mainly existed, three points were chosen. Only one point was measured in the range x < 0 where C. elegans did not often exist.
- Remove the lid of the agar plate, and push the narrow end of a Pasteur pipette with a dropper bulb against the agar and extract the agar plug to make a hole of ø 1-2 mm in the agar layer. Then close the lid.
- From the back of the plate (i.e., the opposite side of the lid), make a hole with a pin vise with a 1 mm drill at the position of the agar hole. This will result in a hole in the same position on the plate and the agar.
- If necessary, use cellophane tape to cover the hole from the back side. Turn back one side of the cellophane tape for ease of peeling. This is not necessary for odorants with large molecular weights such as 2-nonanone. We compared outcomes with and without taping, and found no difference.
- Spot the liquid odorant at the odor source position (we used 2 μl of 30% 2-nonanone diluted in EtOH), immediately cover the lid, place the plate upside-down (hole up) and leave it on the bench.
Note: In the worm’s odor avoidance assay, worms suspended in a small amount of buffer droplet are spotted at the center of the plate 1.5 min before the odor is spotted (Kimura et al., 2010), and the time of odor spotting is counted as t = 0. In this odor measurement, however, the worms are not spotted for the sake of simplicity. - When an appropriate amount of time (i.e., 1, 3, 6, 9, or 12 min) has passed, remove the tape (if applied) without moving the plate. Carefully insert the replacement needle attached to the plastic 2.0 ml disposable syringe so that the needle hole is positioned in the gas phase 1 mm away from the agar surface (i.e., just below the agar surface in the upside-down plate). Slowly extract 0.2 ml of the gas so as not to disturb the gradient severely. Quickly remove the needle tip from the plate and insert it in the gas inlet of the GC, and inject the gas.
Note: Since sampling may destroy the gradient, only one sample was taken from each plate. - Several samples should be taken (we took 7-9) for each position and time, using the median to calculate the dynamic odor gradient.
- On the back side of the agar plate, mark the positions of odor spot and gas sampling with a pen.
- Fitting the odor gradient
- Model selection
At the beginning of curve fitting, a parametric function needs to be specified for the data. Physical phenomena caused in the assay plate are evaporation of the 2-nonanone-ethanol-mixed solution (30:70, v/v) and diffusion of their gaseous molecules in the three-dimensional closed cylindrical space. Although we previously calculated the evaporation and distribution of 2-nonanone numerically (Yamazoe-Umemoto et al., 2015), in the recent study we employed a phenomenological curve fitting to the measured concentration by least squares method for better understanding of the odor gradient (Tanimoto et al., 2017). After the odor sources are put at the two spots, the 2-nonanone concentration C(t) in the plate increases from zero and asymptotically approaches a constant value over time. In this work, a saturation curve C(t) = a(1-exp(-bt)) was used for fitting, where a and b denote an asymptotic concentration and an increasing rate, respectively. This function is a solution of the rate equation dC(t)/dt = b(a - C(t)) which implies that C(t) changes with the rate proportional to the difference from the asymptotic concentration.
In the measurement, increasing of the 2-nonanone concentration was slow as the distance from the spots is far (Figure 7). A mass transfer by molecular diffusion accounts for this result. Therefore the increasing rate is a decreasing function of the distance r from the spot such as b(r) = b0 exp(-b1 r - b2 r2), where b0 (> 0), b1 and b2 are constant parameters. For good fitting in a relatively short time after putting the odor sources, furthermore, the asymptotic concentration is also a decreasing function of r such as a(r) = a0 exp(-a1 r - a2 r2), where a0 (> 0), a1 and a2 are constant parameters. Because there are two odor sources in the plate, the measured concentrations are fitted to the following function with two saturation curves.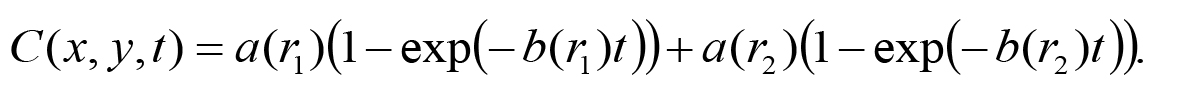
where r1 and r2 are the distances from the position (x, y) on the agar to the two spots (X1, Y1) = (-22, 15) and (X2, Y2) = (-22, -15), respectively.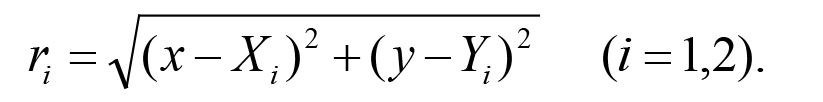
The radius of the plate is 44 mm (1 mm in the plate thickness).
Two constraint conditions are imposed on the fitting. The first constraint condition is that a(r) and b(r) should be decreasing functions of r at least in the range 0 ≤ r ≤ r0, where r0 is the distance from the spot to the edge of the plate (44, 0). From da(r)/dr = - a0(a1 + 2a2 r) exp(-a1 r - a2 r2) and db(r)/dr = - b0(b1 + 2b2 r) exp(-b1 r - b2 r2), inequality conditions da(r0)/dr ≤ 0 and db(r0)/dr ≤ 0 (decreasing even at r = r0) are expressed as: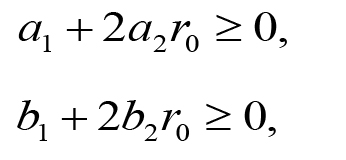
under a0 > 0 and b0 > 0. The second constraint condition is that the asymptotic concentration on the odor sources should be lower than the saturation concentration 34.5 μM of the 2-nonanone (Yamazoe-Umemoto et al., 2015). Letting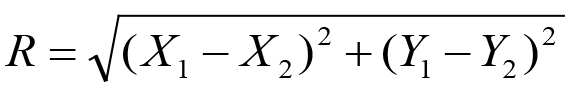 be the distance between the two spots, this condition is expressed as:
be the distance between the two spots, this condition is expressed as: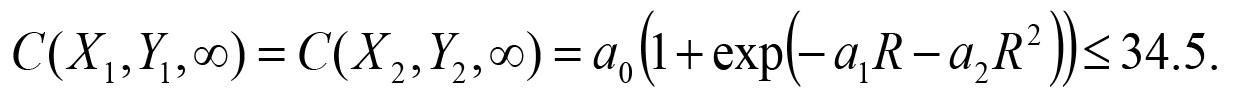
- Fitting algorithm
The fitting parameters in C(x, y, t) are determined by the Levenberg-Marquardt method which is widely used to solve non-linear minimization problems (Press et al., 1992). Letting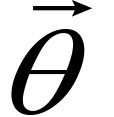 be the parameter vector,
be the parameter vector,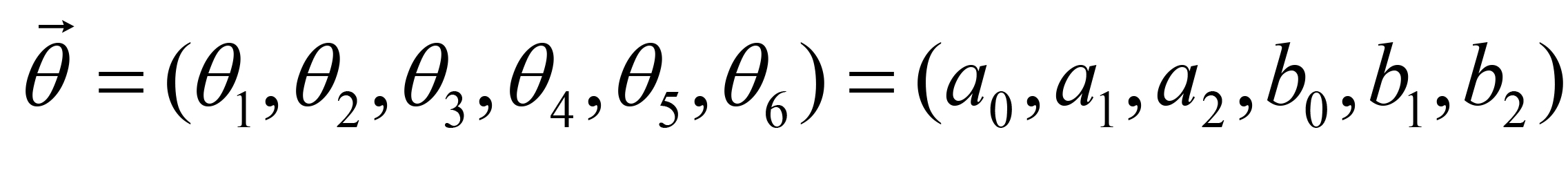
the sum of the squared errors is explicitly defined by: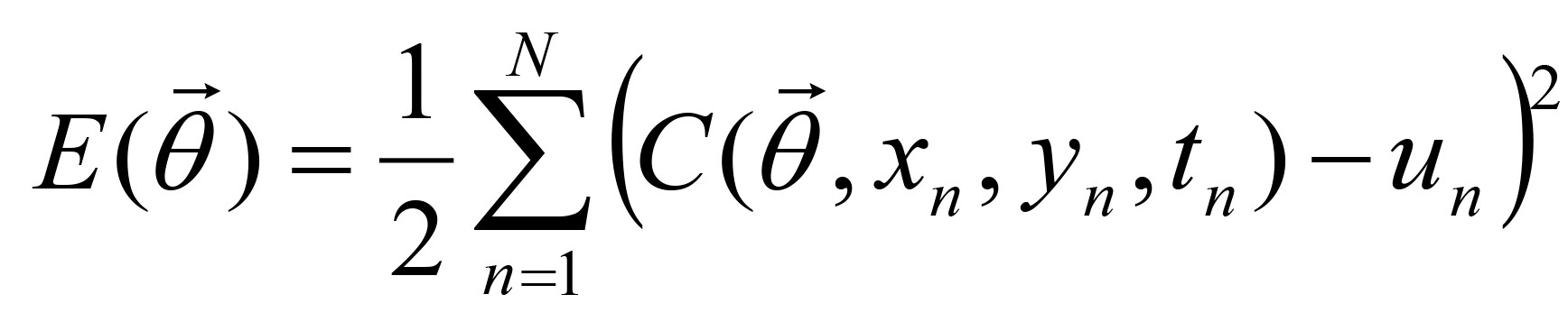
Where un is the n-th measured concentration at position (xn, yn) at time tn ( ). The inequality constraints are expressed as:
). The inequality constraints are expressed as: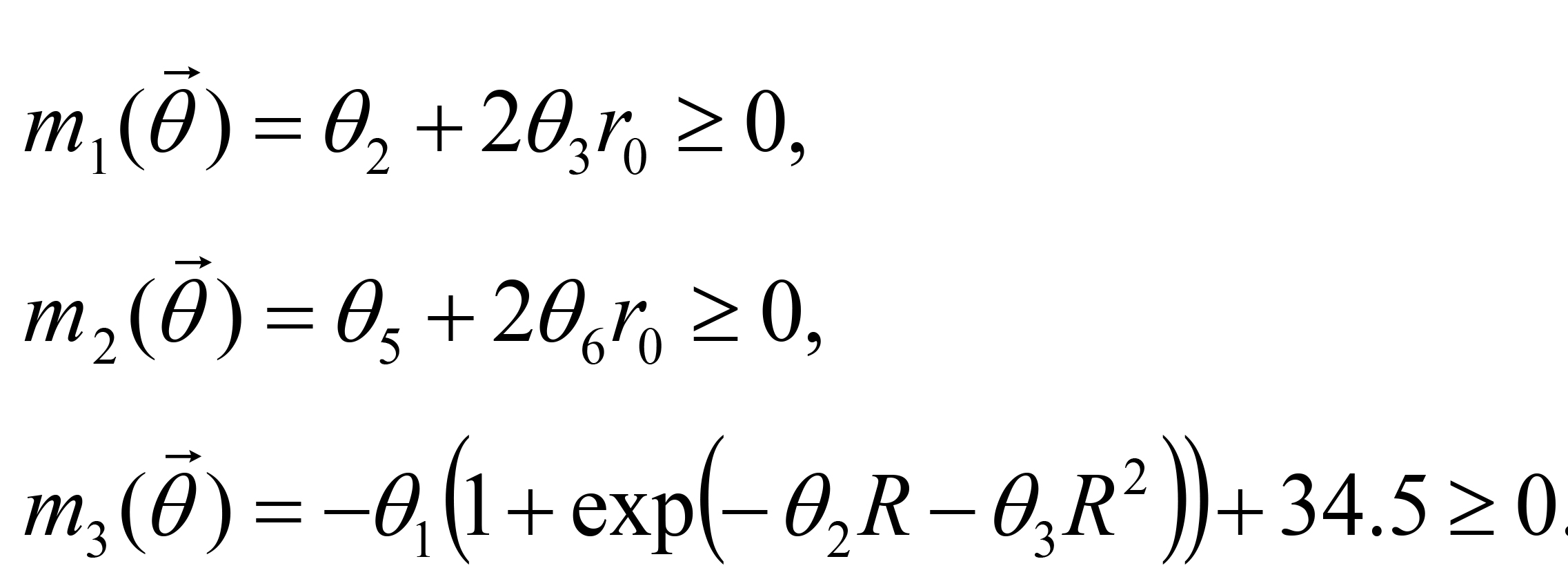
Introducing the following function with logarithmic barriers,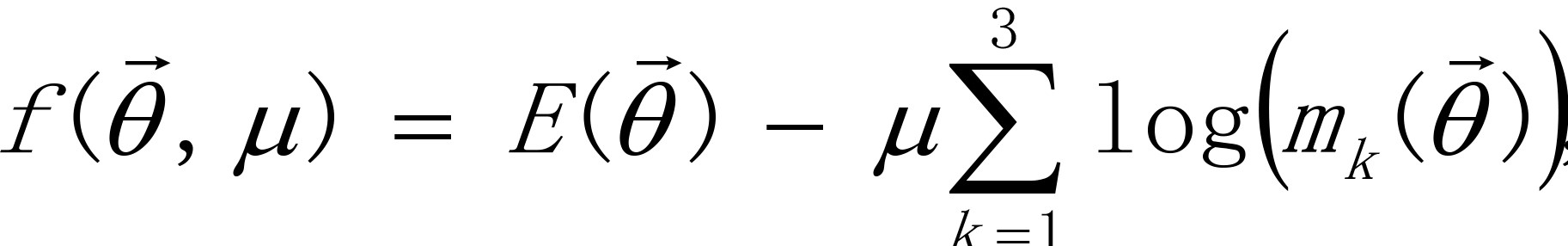
the given constrained minimization problem is approximately replaced by an unconstrained minimization problem. Where μ is a penalty factor whose value is initially large positive and is reduced to zero as is converged. The iterative algorithm to determine which minimizes 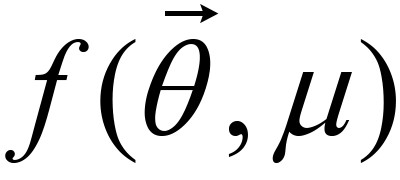 is as follows.
is as follows.
Step 1: Initial values are set for, μ and λ. Where λ is a damping factor in the Levenberg-Marquardt method and is used in Step 2. is chosen to satisfy the inequality constraints. Initial μ and λ are large positive.
Step 2: The Jacobian matrices J = ({Jij}), G = ({Gij}), the diagonal matrix M and the residual vector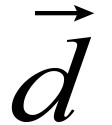 are calculated.
are calculated.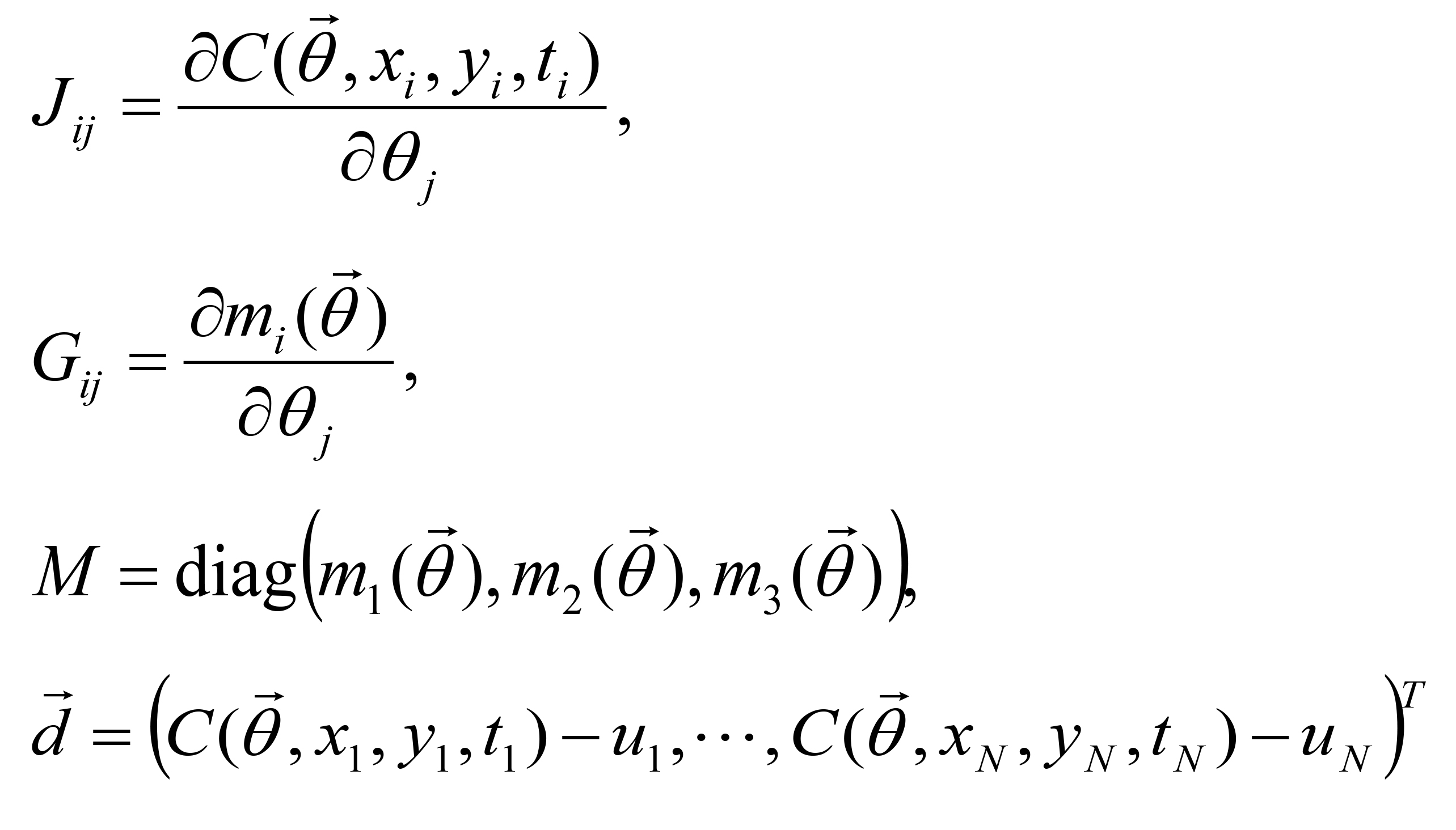
Then, the following linear equation of is solved and
is solved and 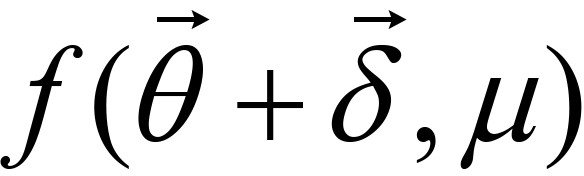 is calculated.
is calculated.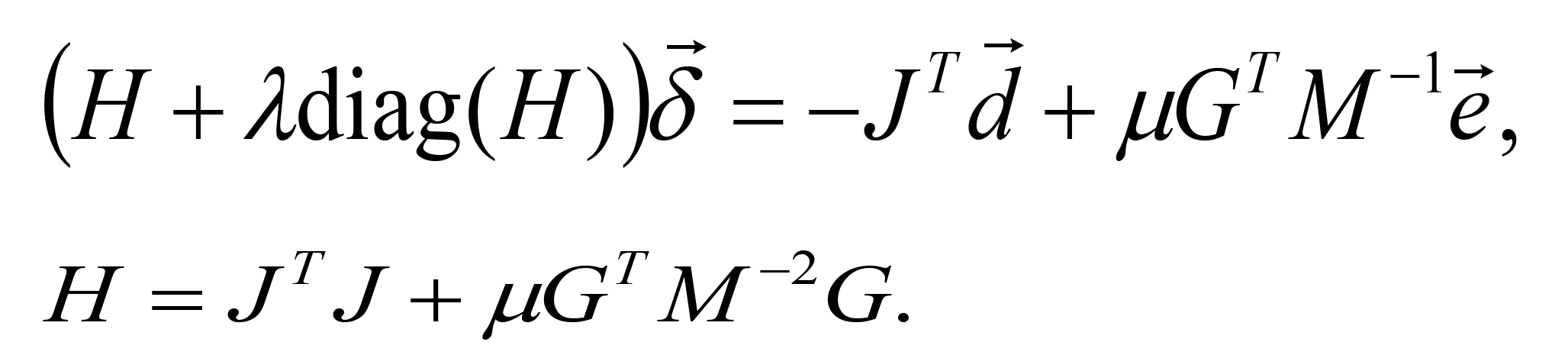
Where
Step 3: If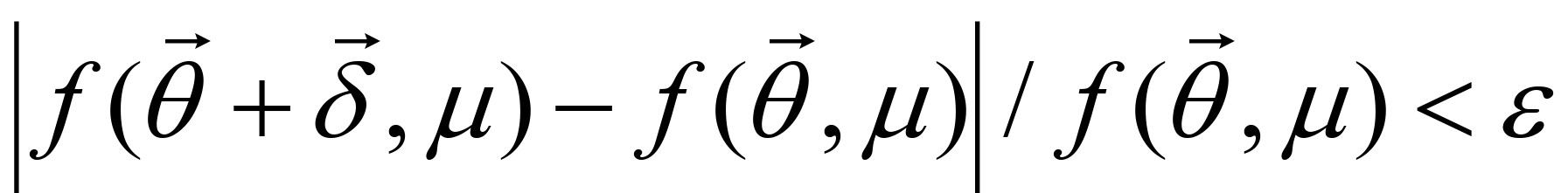 for a small constant ε, then
for a small constant ε, then 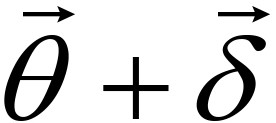 is determined as a solution, or else go to Step 2 with updating , μ and λ. If
is determined as a solution, or else go to Step 2 with updating , μ and λ. If 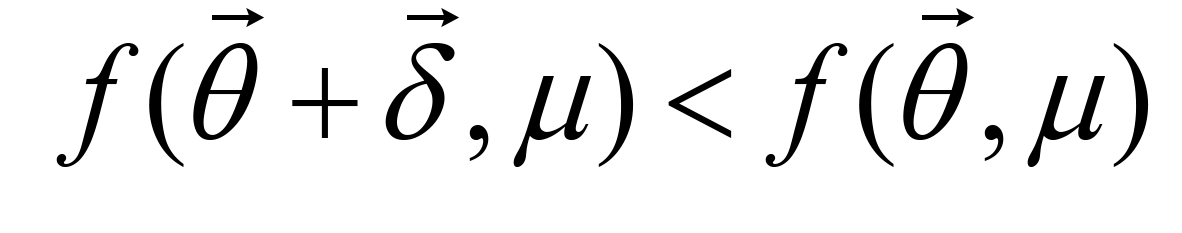 , is updated by checking the inequality constraints. μ and λ are reduced by rates α and β (0 < α, β < 1), respectively. If
, is updated by checking the inequality constraints. μ and λ are reduced by rates α and β (0 < α, β < 1), respectively. If 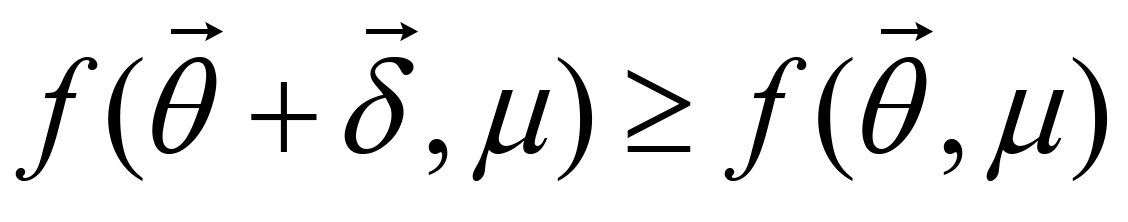 or the inequality constraints are not satisfied, and μ are not updated while λ is increased by a rate 1/β.
or the inequality constraints are not satisfied, and μ are not updated while λ is increased by a rate 1/β. - Execution and result
In this work, the fitting algorithm is implemented in C language and is compiled by the GNU Compiler Collection. The convergence criterion is ε = 1 x 10-6 . Setting of the decrement rates α and β depends on the choice of an initial. In particular, the slow reduction of μ requires the avoidance of invalid updating of , such that the reduction rate α for μ (μ→αμ) is 0.5 < α < 1. When α < 0.5, the penalty factor μ rapidly converges to 0 and an incorrect solution without inequality constraints is derived. Some combinations of initial parameters and decrement rates are tried for fitting. Furthermore, a converged value of is used as an initial value in new iterations with different μ and λ. Good fitting parameters are a0 = 20.68 μM, a1 = 0.7355 cm-1, a2 = -0.05408 cm-2, b0 = 0.8384 min-1, b1 = 0.7835 cm-1 and b2 = -0.05761 cm-2. Some software tools are useful for the non-linear curve fitting. Optimization Toolbox in MATLAB provides packages for non-linear least squares minimization. Solver Add-in in Microsoft Excel is also available for non-linear curve fitting.
Fitting result is shown in Figure 7. Temporal change of the 2-nonanone gradient is shown in Video 1 in Tanimoto et al. (2017). Although the odor sources spread in a round shape (~5 mm in diameter) in the experiment, their shape in the fitting is considered as a point which has no area. Therefore, the fitted 2-nonanone gradient around the spots became pointy.
When a simple exponential function b(r) = b0 exp(-b1 r) was used for fitting, the result was not good. A higher-order correction of more than r2 term requires for good fitting. When b(r) = b0 exp(-b1 r - b2 r2 - b3 r3) was used for fitting, the result was almost the same as that without the r3 term. Fitting using a polynomial function b(r) = b0 - b1 r - b2 r2 - b3 r3 or a fractional function b(r) = 1/(b0 + b1 r + b2 r2) went bad. Fitting using other saturation curve C(t) = c0 t/(t + c1) or C(t) = c0 t2/(t2 + c1) also went bad.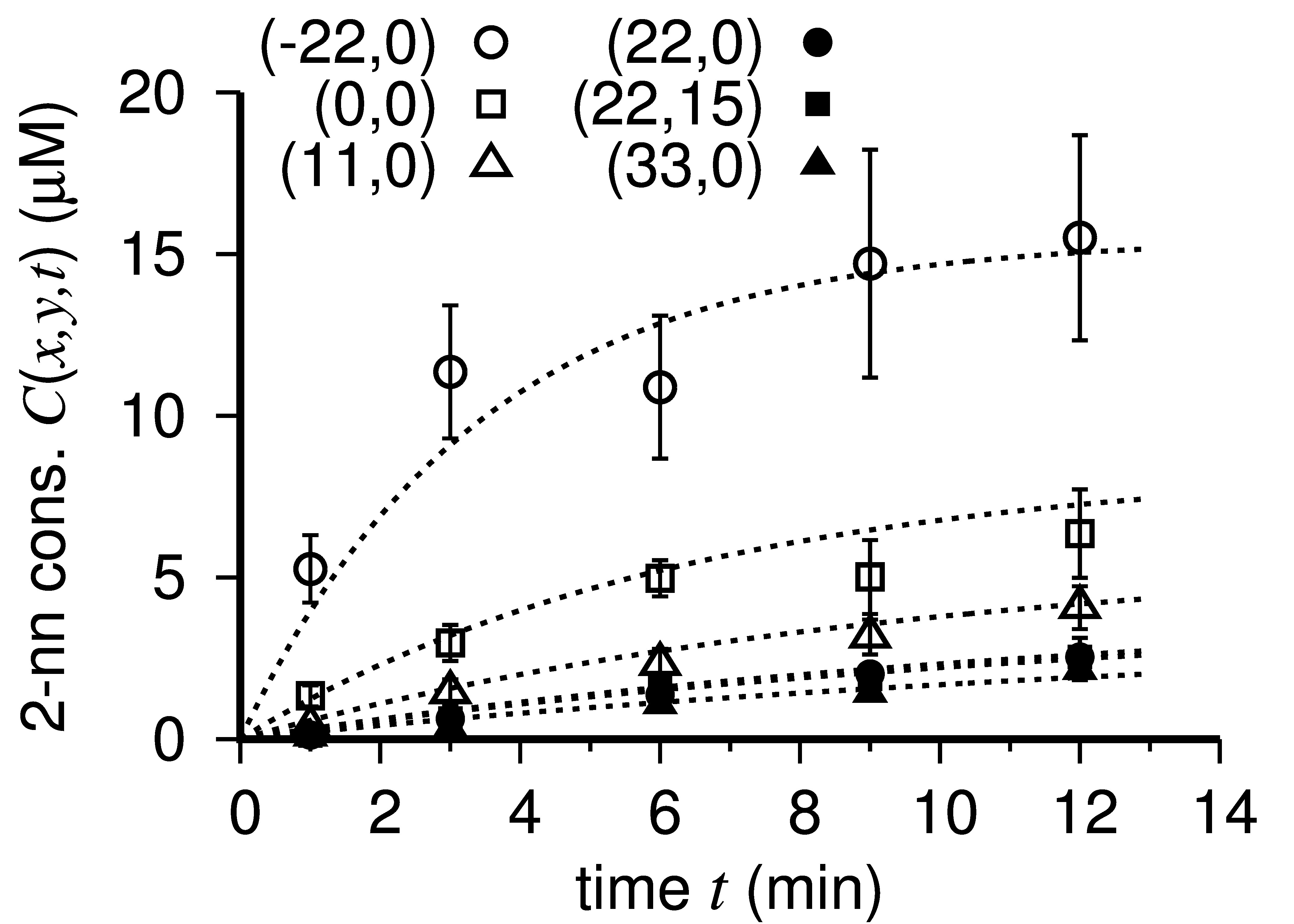
Figure 7. Temporal change of the measured 2-nonanone concentration at the position (x, y) on the agar. The odor sources were put at (-22, 15) and (-22, -15). The broken lines are the fitting curves.
- Model selection
Data analysis
All data related to this study are already published in Tanimoto et al. (2017).
Notes
- We found that variations in odor concentration become considerably smaller with increasing distance from the odor source, i.e., in the right half of the plate. Conversely, variations were greater near the odor source. We consider that this is because diffusion essentially equalizes variation, leading to less variation at greater distances from the source.
- In the natural environment, odors are recognized to exist as plumes; they do not produce a smooth gradient. In this experiment, 0.2 ml was aspirated for one measurement, therefore we were not able to detect any spatial differences within this volume. However, based on the model of the smooth 2-nonanone gradient, C. elegans are estimated to respond to a concentration change of about 0.01 μM/sec, which is consistent with behavioral response in a constant odor concentration change in artificial flow (Tanimoto et al., 2017). This suggests that the gradient is indeed smooth, at least in this case caused by volatilization and diffusion in the static space inside the plastic plate.
- The shape of the gradient can differ substantially depending on the ratio of volatilization to diffusion. In the case of 2-nonanone, the ratio was appropriate for formation of a reasonable gradient.
- The 2-nonanone concentration used in our paradigm is relatively high compared to concentrations of other odorants used in C. elegans odor-taxis analysis, which are in general 10-3-10-4 at odor source (Bargmann et al., 1993). For these odorants, it would be better to use a more sensitive GC. In addition, for these odorants, it may be important to use materials with less adhesion for the vaporizing tank and syringe, such as glass or metal, instead of plastic.
Recipes
- NGM plate (for 1 L)
970 ml ddH2O
3 g NaCl
2.5 g peptone
17 g agar
1 ml cholesterol (5 mg/ml EtOH)
Autoclave; wait until 50-60 °C
Add the following autoclaved buffers:
1 ml 1 M CaCl2
1 ml 1 M MgSO4
25 ml 1 M KPO4 buffer (pH 6) - KPO4 buffer (pH 6.0), 1 M
108.3 g KH2PO4
35.6 g K2HPO4
Add ddH2O up to 1L
Autoclave
Acknowledgments
We especially thank K. Tanaka (FIS Inc., Japan) for all the technical support of odor measurement. This work was supported by a Grant-in-Aid for JSPS fellows (A.Y.-U.), the Osaka University Life Science Young Independent Researcher Support Program, Precursory Research for Embryonic Science and Technology from MEXT, and research grants from Mitsubishi Foundation, Shimadzu Science Foundation, and Takeda Science Foundation (K.D.K.). The authors declare that no competing interests exist.
References
- Bargmann, C. I., Hartwieg, E. and Horvitz, H. R. (1993). Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell 74(3): 515–527.
- De Bono, M. and Maricq, A. V. (2005). Neuronal substrates of complex behaviors in C. elegans. Annu Rev Neurosci 28: 451-501.
- Gershow, M., Berck, M., Mathew, D., Luo, L., Kane, E. A., Carlson, J. R. and Samuel, A. D. (2012). Controlling airborne cues to study small animal navigation. Nat Methods 9(3): 290-296.
- Kimura, K. D., Fujita, K., and Katsura, I. (2010). Enhancement of odor avoidance regulated by dopamine signaling in Caenorhabditis elegans. J Neurosci 30(48): 16365-16375.
- Louis, M., Huber, T., Benton, R., Sakmar, T. P. and Vosshall, L. B. (2008). Bilateral olfactory sensory input enhances chemotaxis behavior. Nat Neurosci 11(2): 187-199.
- Press, W. H., Teukolsky, S. A., Vetterling, W. T. and Flannery, B. P. (1992). Numerical recipes in C. Cambridge University Press.
- Tanimoto, Y., Yamazoe-Umemoto, A., Fujita, K., Kawazoe, Y., Miyanishi, Y., Yamazaki, S. J., Fei, X., Busch, K. E., Gengyo-Ando, K., Nakai, J., Iino, Y., Iwasaki, Y., Hashimoto, K. and Kimura, K. D. (2017). Calcium dynamics regulating the timing of decision-making in C. elegans. Elife 6: e21629.
- Venken, K. J., Simpson, J. H. and Bellen, H. J. (2011). Genetic manipulation of genes and cells in the nervous system of the fruit fly. Neuron 72(2): 202-230.
- Yamazoe-Umemoto, A., Fujita, K., Iino, Y., Iwasaki, Y. and Kimura, K. D. (2015). Modulation of different behavioral components by neuropeptide and dopamine signalings in non-associative odor learning of Caenorhabditis elegans. Neurosci Res 99: 22-33.
Article Information
Copyright
![]() Yamazoe-Umemoto et al. This article is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0).
Yamazoe-Umemoto et al. This article is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0).
How to cite
Readers should cite both the Bio-protocol article and the original research article where this protocol was used:
- Yamazoe-Umemoto, A., Iwasaki, Y. and Kimura, K. D. (2018). Measuring Spatiotemporal Dynamics of Odor Gradient for Small Animals by Gas Chromatography. Bio-protocol 8(7): e2797. DOI: 10.21769/BioProtoc.2797.
- Tanimoto, Y., Yamazoe-Umemoto, A., Fujita, K., Kawazoe, Y., Miyanishi, Y., Yamazaki, S. J., Fei, X., Busch, K. E., Gengyo-Ando, K., Nakai, J., Iino, Y., Iwasaki, Y., Hashimoto, K. and Kimura, K. D. (2017). Calcium dynamics regulating the timing of decision-making in C. elegans. Elife 6: e21629.
Category
Neuroscience > Behavioral neuroscience > Chemotaxis
Biochemistry > Other compound > 2-nonanone
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.