
- 实验方法

- Request a Protocol
- 关于
-

研究背景
计算流体动力学作为计算机科学、流体力学与数值计算方法深度融合的交叉学科,已在航空航天、汽车船舶、能源环境、生物医学、采矿冶金等领域广泛应用。在生命科学领域,计算流体动力学不仅被用于生物医学工程中的血液循环系统模拟和呼吸系统气体流体动力学等分析,更是研究现生生物生理功能与形态适应的重要工具 [1–3]。近年来,随着虚拟-古生物技术发展,计算流体动力学逐渐应用于古生物学领域。通过整合化石形态特征与古环境信息,可对灭绝生物的运动模式、捕食机制及其与环境的交互关系进行研究分析。相较于传统研究手段,计算流体动力学技术可通过建立化石三维复原模型,分析复杂流体与生物体之间的相互作用 [4],为探究古生物生存策略、演化路径及其生态适应性提供量化依据。目前计算流体动力学在古生物领域的应用主要集中于古生代海洋无脊椎动物研究 [4–9],在古脊椎动物研究中的应用相对较少。已有的研究包括对古生代鱼类、三叠纪海生爬行动物 [10,11] 等进行形态功能定量分析与运动模式研究。就古鱼类研究而言,目前研究多采用计算流体动力学模拟求解鱼类游泳参数(如升力、阻力等),研究对象多集中于身体构型较为简单的无颌类 [12,13]。对有颌鱼类的相关研究相对较为匮乏,多为结合现生物种的对比分析 [14]。
计算流体动力学研究方法与流程
1. 基本原理
计算流体动力学通过计算机模拟手段来分析各种流体流动、传热及相关物理问题,而所有流体问题都遵守质量守恒定律、动量守恒定律和热量守恒定律。若不考虑热量交换,大部分流体问题仅关注连续性方程(质量守恒)和纳维-斯托克斯方程(N-S方程,即粘性不可压缩流体动量守恒的运动方程),二者共同描述了流体的质量、动量和能量在空间和时间上的变化规律。连续性方程表示在单位时间内流入系统的流体净质量和该系统在单位时间内的质量增量是相等的,数学表达式如下: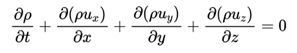
式中,ux,uy,uz分别为x,y,z方向在t时刻的速度分量,t为时间,ρ为流体密度。纳维-斯托克斯方程描述了在粘性不可压缩的流体系统内,系统内所受的外力的总和等于系统内部的动量的变化率,在直角坐标系的表示如下: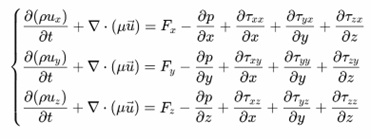
式中,ρ为流体密度,ux,uy,uz分别为x,y,z方向在t时刻的速度分量,τ为应力张量,F为应用在物体上的力,若物体只受重力影响时,Fx = Fy = 0,Fz = ρg。计算流体动力学通过数值方法通过求解N-S方程来模拟流体流动及其与固体边界的相互作用,进而对流速、压力、浮力、涡量等关键参数展开分析。在水生生物游动过程中,雷诺数(Re)是描述流体特性的一个核心无量纲参数,定义为流体惯性力与粘性力的比值,是判别流体流动状态(层流或湍流)的关键指标,表达式为: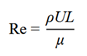
式中,ρ表示流体密度(kg/m3),U为流体速度(m/s),L是特征长度(m),μ是粘性系数(kg/(m·s))。在低雷诺数条件下(通常对应毫米级以下的微小生物),粘性力占主导,流体流动方式是层流,阻力主要由表面摩擦贡献;高雷诺数条件下(通常对应于厘米级及以上尺度的生物),惯性力占主导,流动变为湍流。目前研究关注的脊椎动物类群多为厘米级及以上的宏观尺度,处于高雷诺数环境,故流体动力学分析的重点在于层流状态的维持以及湍流边界层的有效控制。针对湍流模拟,可以大致分为三类计算流体动力学方法:直接数值模拟(DNS)方法、大涡模拟(LES)方法和雷诺平均方法(RANS)方法。在工程领域中较常用的是雷诺平均方法方法,另两种直接数值模拟和大涡模拟方法计算量与成本较高,对计算机性能要求高 [15]。
在化石鱼类的计算流体动力学研究中,鱼体表面存在一个很薄的剪切层,即边界层。该层内流体速度从壁面处零速沿法向迅速增至主流速度,其外的流场可视为无粘性流动(图3A)。边界层的存在使得壁面附近产生较大剪切力,其大小与流体粘度和壁面速度梯度相关。当攻角较小时,鱼体表面边界层保持较薄的附着状态;当攻角增加到某一临界值时,边界层在逆压梯度与粘性的共同作用下发生分离,流体卷起形成旋涡,主流远离壁面,形成低速的有回流的分离区(图3B)。边界层分离会显著增加流动阻力与和流动能量损失,对于鱼体游动等绝热流动,损失来源于粘性耗散。
2. 软件选择
计算流体动力学研究工具主要分为商业软件(如ANSYS Fluent, Autodesk CFD, COMSOL Multiphysics等)与开源软件(如Fluidity, OpenFOAM, SU2, OpenLB等)两大类别(表1)。商业软件中,ANSYS Fluent具备丰富的物理模型与求解器,可精准解析古生物在流体环境中的运动特征;Autodesk CFD凭借高集成度优势,便于实现设计流程与流体分析的无缝整合;COMSOL Multiphysics则具有强大的多物理场耦合能力,适用于化石鱼类运动与环境相互作用的复杂系统建模。开源软件中,Fluidity基于有限元方法,在大规模科学计算领域展现出高效并行计算性能;OpenFOAM作为高度灵活的开源工具包,支持自定义求解器与算法开发,可以满足前沿科研需求 [16];SU2则专注于航空航天领域的气动分析与优化,为流体动力学研究提供多元算法支持。
在数值计算方法层面,最常用的计算流体动力学算法包括有限体积法(FVM)、有限差分法(FDM)和有限元法(FEM),除此之外还有粒子法,如光滑粒子法(SPH)、半隐式运动粒子法(MPS)、有限体积粒子法(FVP),其中光滑粒子法方法计算速度较快,但准确度不如更适用于工程场景的半隐式运动粒子法和有限体积粒子法。近年来新兴的格子玻尔兹曼法(LBM)则是通过模拟流体粒子在离散网格上的运动和碰撞来捕捉宏观流体行为,因其在处理复杂几何边界和并行计算效率方面的潜力而受到关注,在生物流体力学和极端环境模拟等前沿领域中有所运用 [17]。采用该法的软件有OpenLB, Palabos, SIMULIA XFlow, Altair ultra FluidX, SIMULIA PowerFLOW, TF-Lattice等,部分商业软件中也集成了LBM插件,但该方法的计算量随分辨率的提高呈指数增长,需要依赖超算实现大规模并行计算。总而言之,古生物领域计算流体动力学相关研究大多采用ANSYS Fluent和COMSOL Multiphysics进行数值模拟与流体力学分析。为进行更高精度、数据规模更庞大的计算,探索更加复杂的生物-流体相互作用机制,研究者可以尝试运用开源软件如OpenFOAM [16],结合格子玻尔兹曼法、大涡模拟 [18] 等新兴方法来计算。
表1. 计算流体动力学主流仿真软件方法比较与总结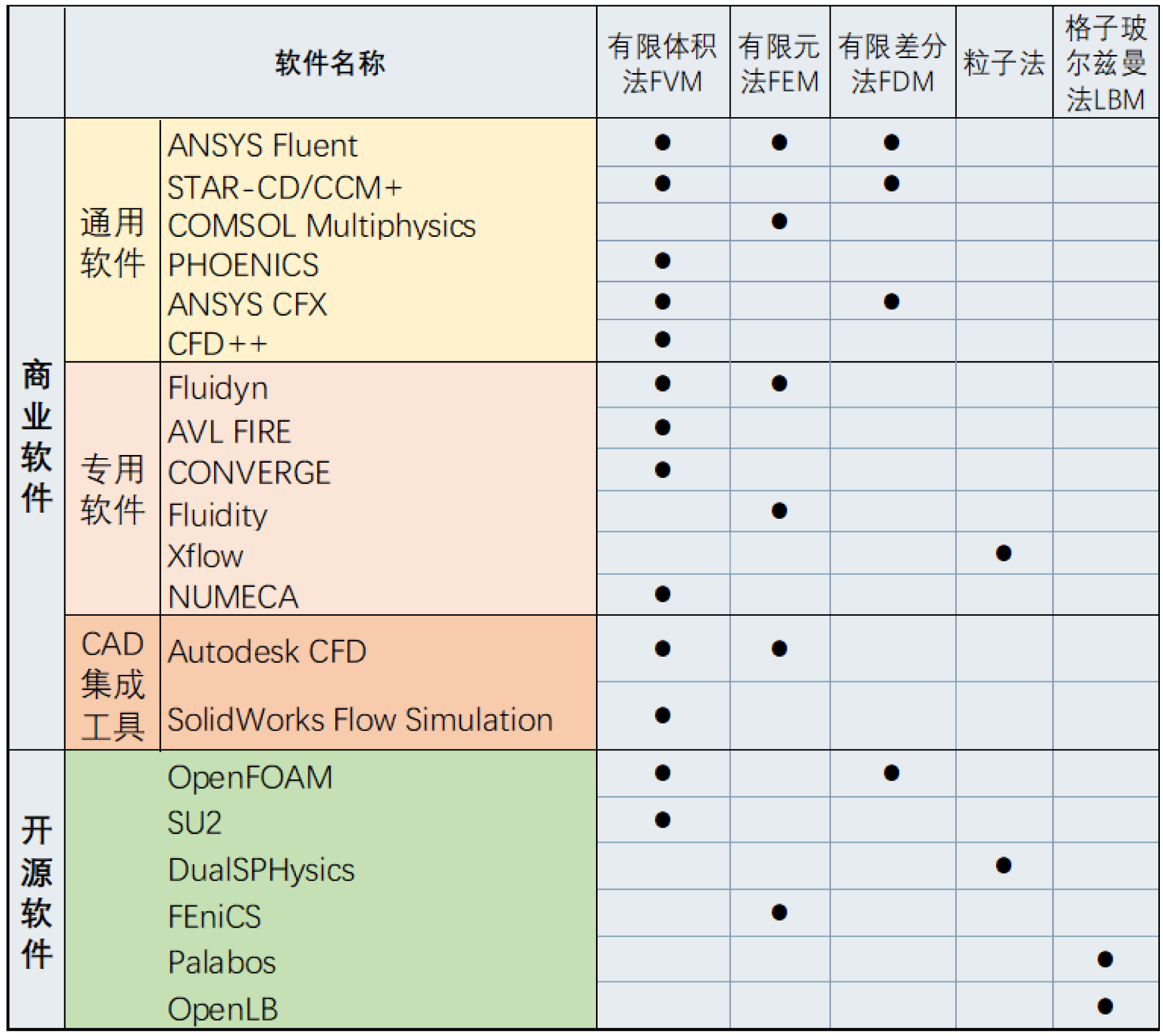
3. 研究流程
a. 模型构建与预处理:基于化石CT扫描数据建立高精度三维数字模型,复原由于保存原因产生错位、变形或缺失的化石结构。为满足后续流体动力学数值模拟的需求,将原始模型数据(如OBJ或STL格式)转化为ANSYS Fluent或COMSOL能够识别的高精度几何格式,如IGES(.igs)或Parasolid(.x_t)。软件选择:Blender,Rhinoceros,ZBrush,Mimics,VG Studio,SolidWorks,VSG Avizo。
b. 计算域构建:三维化石模型导入(CT扫描/激光扫描数据);并围绕模型建立流域(长方体或圆柱体,通常上游延伸3倍体长,下游10倍体长,以确保流场充分发展)。软件选择:ANSYS Spaceclaim,SolidWorks。
c. 网格划分:自动或手动生成网格(二维面网格采用三角面片或四边形面片;三维体网格采用四面体或六面体单元);在流-固界面添加棱柱层,同时重点区域加密以捕捉边界层效应,根据具体需求进一步优化网格数量与质量。软件选择:snappyHexMesh,3matics,ANSYS Meshing,Fluent Meshing。
d. 参数设置:指定流体属性(密度、粘度),选择层流或湍流模型(如k-ε/k-ω/RANS/SST等模型),设置边界条件(滑移/无滑移边界,入口流速,出口零压等)。软件选择:ANSYS Fluent, COMSOL Multiphysics。
e. 模拟求解:稳态/瞬态求解,对比实验数据(如水槽或风洞实验)验证模型可靠性。软件选择:ANSYS Fluent,CFX,COMSOL Multiphysics。
f. 结果后处理:计算阻力系数(CD)、升力系数(CL)等参数,可视化流场(绘制流线图/流速云图/颗粒追踪图等),利用结果分析研究对象的流体力学性能、摄食效率、运动方式等。软件选择:ANSYS Fluent,CFD-Post,COMSOL Multiphysics。
4. 结果与分析
计算流体动力学模拟的结果通常涵盖灭绝生物的水动力性能、运动特性及捕食方式等多维度分析,可以通过流速图、压力分布图等不同方式进行可视化。以古鱼类为例,通过在不同流速与攻角条件下测量鱼体所受阻力(FD)与升力(FL)、表面压力分布、升/阻系数以及升阻比等参数,可量化评估古鱼类的推进效率、运动稳定性与机动性。
鱼类在游动过程中,流体对鱼体施加壁面剪切应力和压力应力,升力则是二者在垂直于流动方向的合力,阻力为平行于流动方向的合力。鱼体所受阻力大致可划分为压差阻力、摩擦阻力,分别源于鱼体前后压力差、粘性流体与鱼体表产生剪切应力,二者大小取决于鱼体几何形状与流动方向的关系。鱼体所受升力源于上下表面压力差,与鱼体形态、攻角(α,纵轴与来流之间的夹角)、流体速度与密度等因素相关。阻力系数(CD)和升力系数(CL)的数学表达式为: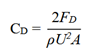
式中,ρ表示流体的密度,U为入口速度,A指模型的表面积,升阻系数均无量纲,对于鱼体等流线型物体,阻力系数随着雷诺数的增加而逐渐减小,而当层流向湍流转捩后,阻力系数开始增加;升力系数一般随攻角增加而增加,到达临界点后降低并产生失速现象。与机翼的空气动力学研究相似,鱼类游泳的流体动力学研究关注如何提升水动力性能与游动效率——即减小阻力,增大推动力。现生游泳动物减少阻力的两种机制是:保持层流附着以减少摩擦阻力,和湍流延迟边界层分离以减少压力阻力。计算流体动力学模拟揭示了古鱼类通过其流线型体态、尾鳍形态、湍流控制结构(如鳍、鳃等),以及改善鳞片、皮肤等表面摩擦等策略降低流体阻力的演化机制 [19]。水生动物提升流体动力学性能方法大致有被动减阻机制(被动结构优化如楯鳞表面的平行脊状结构、皮肤分泌物、鳍状肢结节等)、主动减阻机制(主动运动控制水流如尾鳍推进、背/臀鳍调节、鳍条结构等)以及环境涡流利用(如鱼类集群、卡门步态运动等)[20]。在生态位研究中,底栖物种可利用地面效应增强升力,从而获得比浮游或游泳更高的升阻比 [13];而远洋物种则多依赖流线型体态减小阻力。
古生物学领域中计算流体动力学的应用
计算流体动力学技术在工程与生物医学领域已形成较成熟的应用体系 [21–23],而在古生物学研究中尚处于发展阶段,目前主要聚焦于不同化石类群在流体环境中的形态功能与运动学分析(表1),尤其是古生代海洋无脊椎动物的流体模拟分析 [24]。计算流体动力学是古生物形态功能定量分析的重要工具,通过模拟能够分析不同灭绝物种在海洋、湖泊、河流等流体环境中的运动方式(如底栖、游泳、浮游等),以及流体动力条件对其形态结构、生活方式和演化的影响,为古生物功能形态学研究开拓了新的视野与研究思路。
表2. 计算流体动力学技术在古生物学领域中的研究实例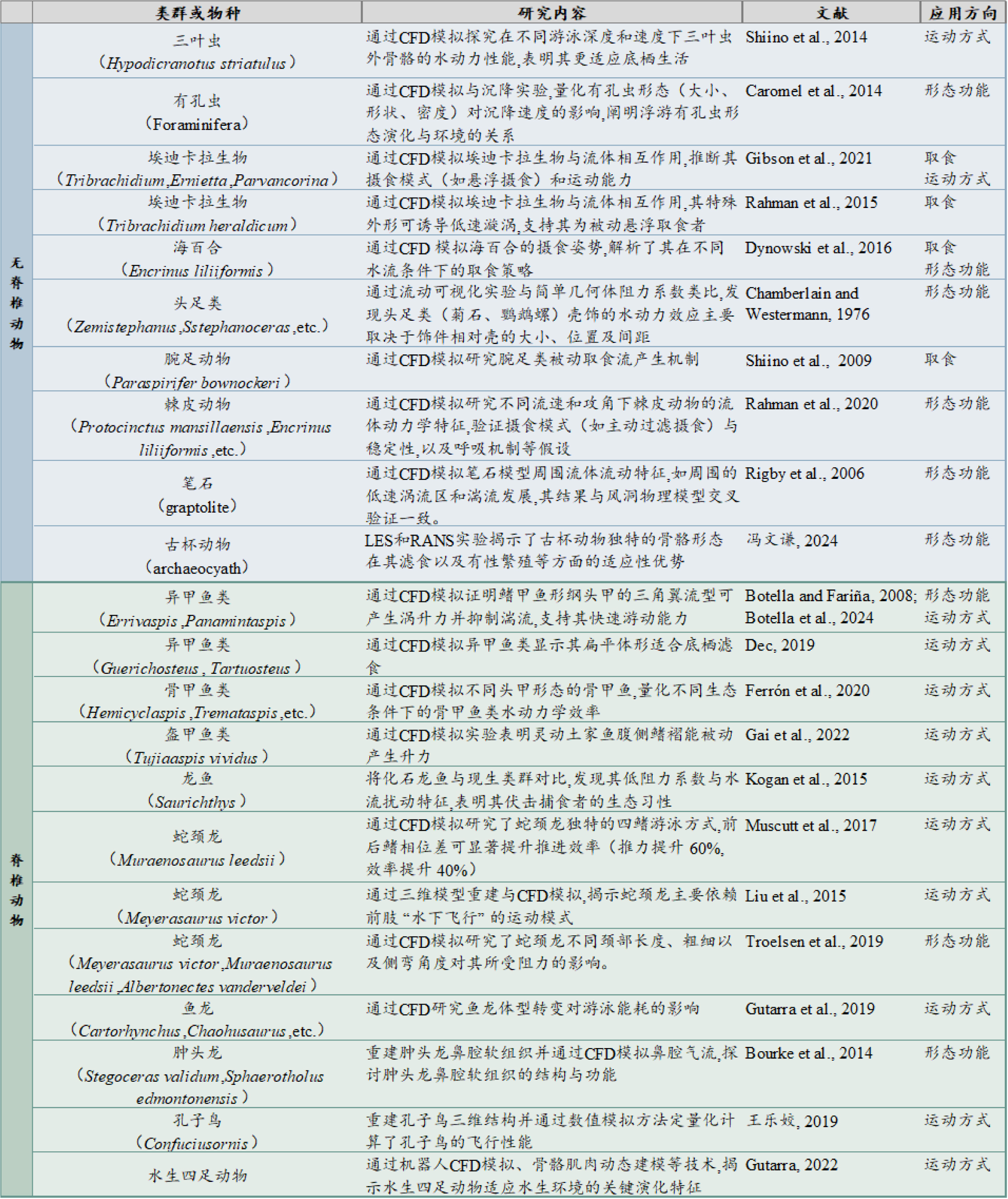
无脊椎动物(图1)
计算流体动力学技术在海洋无脊椎动物研究中应用较多,揭示了多个类群的流体动力学适应性特征。Shiino等通过计算流体动力学模拟了三叶虫(Hypodicranotus striatulus)在不同游泳深度和速度下其外骨骼的水动力性能,表明其更适应底栖生活 [6]。Caromel等通过对有孔虫进行计算流体动力学模拟研究其形态(大小、形状、密度)对沉降速度的影响,阐明浮游有孔虫形态演化与环境的关系 [7]。此外,Rahman等对埃迪卡拉纪生物(Tribrachidium heraldicum)进行计算流体动力学研究,发现其特殊外形可诱导低速漩涡,支持其为被动悬浮取食者的观点,将悬浮取食的证据提前到5.55–5.50亿年前 [8]。Dynowski等通过计算流体动力学模拟海百合(Encrinus liliiformis)的摄食姿势,解析了其在不同水流条件下的取食策略 [9]。其他研究还包括头足类贝壳纹饰的流体动力学功能 [25],及腕足动物被动取食流产生机制 [5]。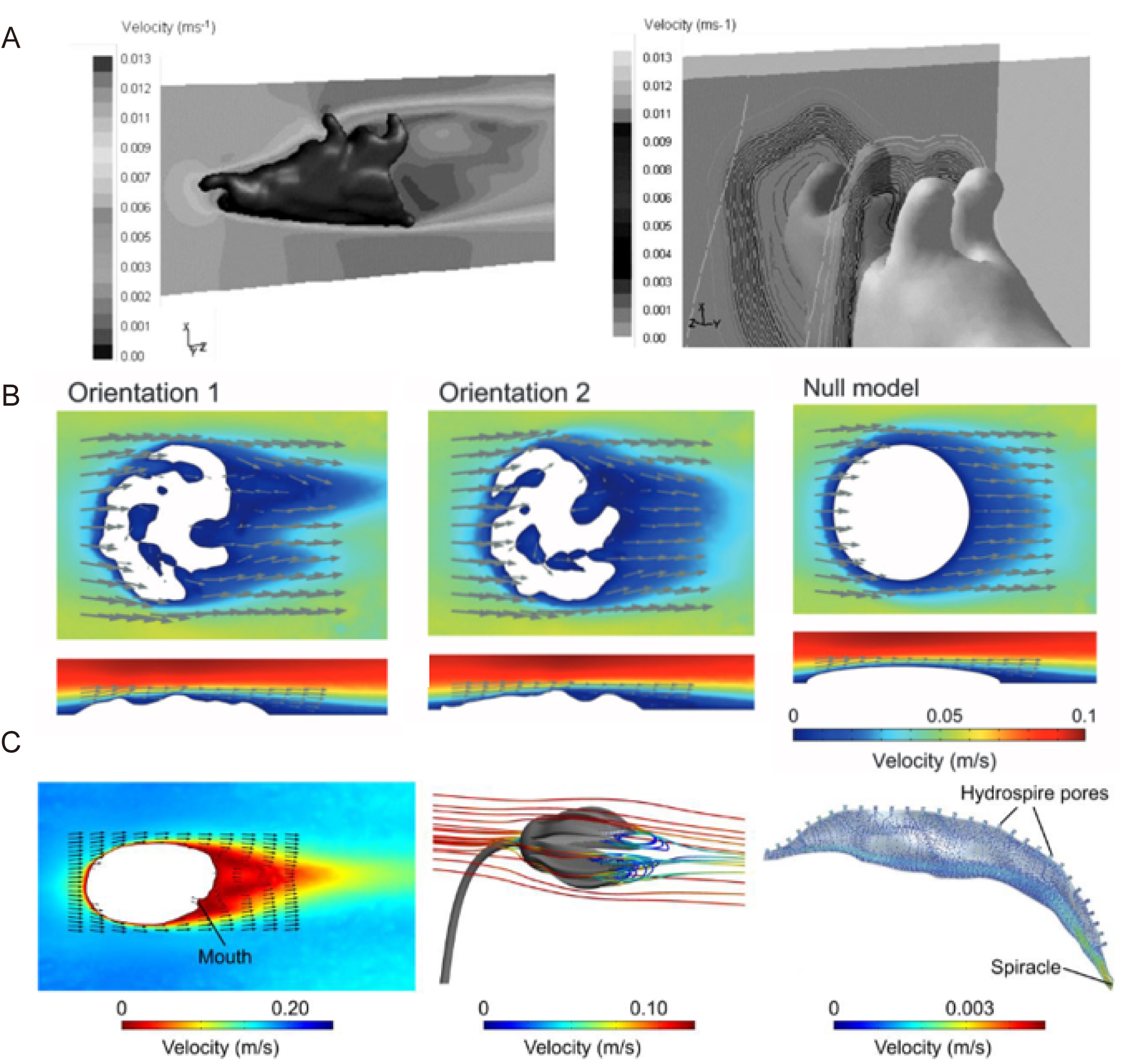
图1. 部分化石无脊椎动物计算流体动力学应用与分析案例。A,对笔石模型计算流体动力学模拟的三维与横截面速度云图 [48]。 B,埃迪卡拉生物(Tribrachidium)在不同攻角情况下的流速示意图[8]。C,水流绕海百合和海蕾模型的流速与流线图 [20]。
脊椎动物(图2)
在脊椎动物研究中,计算流体动力学模拟在无颌类、有颌鱼类及水生四足动物的运动力学分析中有所运用。对无颌类的研究发现,鳍甲鱼类头甲的三角翼流型可产生涡升力并抑制湍流,支持其快速游动能力 [12,26]。Ferrón等则通过对比骨甲鱼头甲形态,量化了底栖或远洋等不同生态条件下鱼体的水动力学效率 [13]。Dec对异甲鱼类(Psammosteids)的模拟显示其扁平体形适合底栖滤食 [27]。Gai等对灵动土家鱼(Tujiaaspis vividus)的计算流体动力学实验表明其腹侧鳍褶能被动产生升力 [28]。
在有颌鱼类研究中,Kogan等将化石龙鱼与现生类群对比,发现其低阻力系数与水流扰动特征,表明其伏击捕食者的生态习性 [14]。在水生四足动物研究中,Gutarra等结合解剖学与物理建模方法,通过计算流体动力学、机器人模拟、骨骼肌肉动态建模等技术,对次生水生四足动物的运动机制研究,揭示其适应水生环境的关键演化特征,如流线型体型降低阻力、鳍状肢推进效率、浮力控制及运动模式 [29]。关于蛇颈龙类的相关研究较多,Troelsen等对具有不同颈长与颈粗的蛇颈龙进行计算流体动力学模拟,发现颈部长度对直线游动阻力无显著影响而较粗颈部可降低阻力,推测其长颈演化可能与扩大捕食范围而非流体力学优势相关 [30]。Muscutt等对蛇颈龙鳍肢协同运动的计算流体动力学模拟显示,前后鳍相位差可显著提升推进效率(推力提升60%,效率提升40%)[31]。Liu等通过三维模型重建,揭示一种蛇颈龙(Meyerasaurus victor)主要依赖前肢驱动游泳,后肢在推进中作用较小,主要功能为辅助转向和稳定,整体为“水下飞行” 的运动模式 [11]。除蛇颈龙外,计算流体动力学模拟还用于鱼龙身体形态研究,如Gutarra等对9种鱼龙和宽吻海豚的三维数字模型进行水流模拟,首次量化了身体形态转变对鱼龙游泳能量需求的影响 [10]。
此外,计算流体动力学还用于空气动力学相关研究,如肿头龙类(Pachycephalosauridae)鼻腔气流功能研究 [32] 以及孔子鸟(Confuciusornis)飞行性能探究 [33] 等,但古鸟类飞行性能的空气动力学相关研究较少,可能是未来潜在的重点研究方向。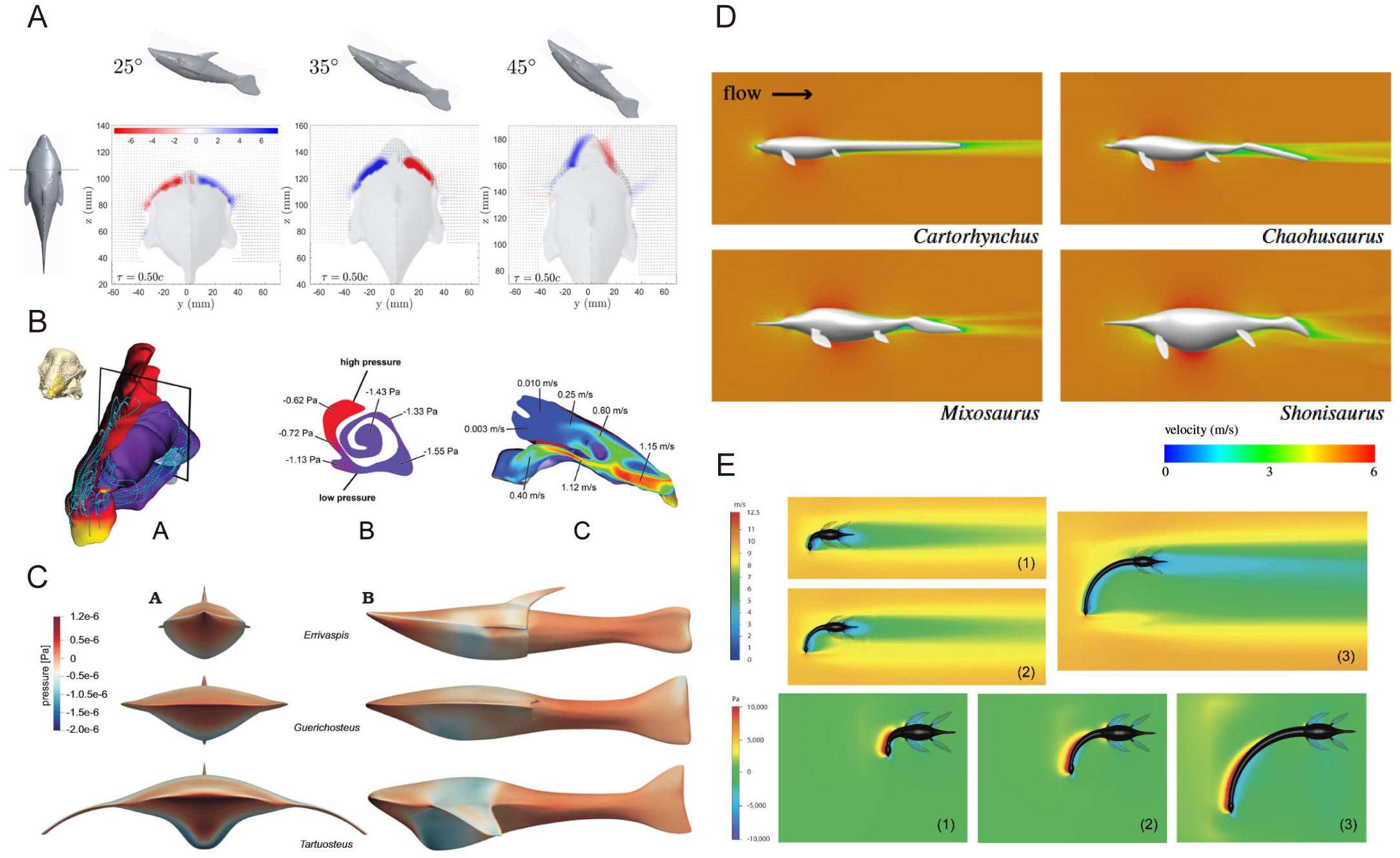
图2 部分化石脊椎动物的计算流体动力学应用与分析案例。A,异甲鱼(Panamintaspis snowi)在不同攻角情况下头甲附近的无量纲涡量图 [26]。B,肿头龙(Stegoceras validum)鼻甲模型与鼻腔内气流压力和速度分布 [32]。C,流速为1.5m/s时异甲类鱼体压力分布图 [27]。D,流速为1.5m/s时鱼龙周围水流速度二维示意图 [10]。E,蛇颈龙在不同颈长、不同水流速度条件下的流速和压力分布图 [30]。
总结及展望
1. 计算流体动力学应用局限性
古生物硬体结构常因压实作用与矿物交代作用而缺失、变形或异位,肌肉及器官等软组织结构则通常难以在化石记录中完整保存。肌肉分布与皮肤弹性等软组织特征对部分生物游泳推进效率的贡献率可达30% [4],同时,古生物的运动方式直接影响流体动力学特征,但化石类群难以单纯从形态学特点进行推断行为特征,故形态数据和运动数据的缺失制约了流体动力学重建模型的精度和求解计算的可靠性。
由于与古生物相关的古环境数据欠缺,多数仿真研究将模拟海床的壁底设置为光滑表面的理想情况 [13,26],而未考虑底质类型与粗糙度对边界层流动的影响,但这对于底栖鱼类运动模拟尤为关键。同时,水温、水流速度等生态因子通过改变流体黏度与雷诺数显著影响鱼体水动力性能,然而古环境数据的不确定性增大了模型误差。
由此可见,仅依赖形态学特征难以全面研究化石类群的流体动力学特性,需结合现生类群的研究成果,才能更准确地推测化石鱼类的运动能力。如关于化石水生四足动物的研究结合了现生半水生动物(如海狮、水獭)的运动学数据作为功能类比,并参考了化石骨骼形态与软组织痕迹(如皮肤印痕、鳞片排列)进行复原 [29]。
2. 未来研究方向
计算流体动力学技术在古生物研究中具有多重优势,相比风洞或水槽实验而言,计算流体力学方法成本较低,更为快捷灵活,其非破坏性的分析避免了对脆弱化石样本的物理损伤,且支持复杂模型多参数测试。但计算流体动力学在古生物领域中仍具局限性,如缺乏可用于跨类群对比分析的标准化形态数据库与开源几何模型;在复杂水动力环境下湍流模型的选择与边界层处理等问题仍存在争议 [20];缺乏灭绝类群的实验数据来辅助验证计算模拟结果等,故该方向仍有广阔的提升空间与发展潜力。未来化石鱼类研究需要结合古生物学、形态学、比较解剖学、计算流体力学、生物力学、仿生学及生态学等多领域理论与方法,进行多学科交叉合作的同时也加强与现生鱼类的形态学比较研究,探讨不同类群演化过程中游泳模式和形态功能趋同的流体动力学特征变化规律,为揭示生物与环境共演化的机制提供更全面的科学依据。以古鱼类为例,未来研究可集中于以下方向:
2.1 三维动态分析
化石鱼类尚未建立系统的推进模式研究框架,未来可以借鉴现生鱼类游动的两种推进模式:身体/尾鳍(BCF, Body/Caudal Fin)和奇鳍/胸鳍(MPF, Median/Pectoral Fin)[34–36],根据鱼类形体结构、运动特性等又可以将BCF分为鳗鲡模式、尾鳍摆动模式、鲹科模式,根据胸鳍运动方式将奇鳍/胸鳍推进分为前后排翼运动、摇翼运动、上下拍翼运动三类 [37]。身体/尾鳍推进在快速、高效、连续运动中具明显优势,而奇鳍/胸鳍推进更为平稳、机动,在低速运动和转弯中占优,二者在现生鱼类和仿生鱼机器人推进方式的研究中多有讨论 [38,39]。但目前古生物开展的流体力学模拟多为静态条件 [12,13,26],无法模拟生物的动态运动(如摆尾、振翅、附肢摆动等)。仿生流体力学领域多运用动网格模拟现生鱼类摆尾游动,对于化石鱼类只能依赖于现生鱼类波模型 [40],并综合二维或三维动网格技术来探究摆动频率、幅度对于游泳速率的影响。
2.2跨类群与群体分析
生物通过集群运动通常能获得空气动力学或流体力学优势,如鱼群集群游动、鸟类编队飞行等。目前古生物领域的群体流体动力学分析仅在无脊椎动物如埃迪卡拉生物中有所应用 [41],脊椎动物大多研究仅进行了单模型模拟,未来可以尝试结合沉积学数据(如古水流方向与底质沉积物分布等),进行多模型模拟群体流体力学分析,研究古生物群落的运动或摄食模式。借鉴现生鱼类集群行为研究 [42,43],未来可以探究古鱼类集群游动的的编队形态、协同交互机制,对比下游个体与上游个体所受阻力大小,揭示其群体行为的流体动力学优势。
除了目前常见的单物种或单类群分析外,也可以进行跨类群功能演化分析,通过对比不同无颌鱼类或有颌鱼类化石类群的流体力学特征,揭示形态适应性演化路径。目前多集中于无颌的骨甲鱼类 [13]和异甲鱼类[26,27] 的流体力学分析,而盔甲鱼类头甲形态演化与其他有颌类如盾皮鱼、棘鱼等形态演化研究目前相对空白。
2.3 多方法技术综合研究
计算流体动力学可以结合其他技术方法对复杂问题进行分析,如有限元分析(Finite element analysis,FEA)[44]、多体动力学分析(Multibody dynamics analysis,MDA)[45]、几何形态测量学(Geometric morphometrics,GMM)[13] 等,重点关注有软组织痕迹留存的化石,综合肌肉与软组织重建结果来进行运动学分析,更精准地研究古生物化石与流体环境的流固耦合问题。目前古鱼类计算流体动力学模拟并没有重建肌肉或软组织的研究实例,未来可以借鉴其他类群咬合力分析或运动能力研究中的重建肌肉重建方法进行尝试 [46,47],分析特定部位肌肉如尾鳍肌肉等对运动特性的影响等。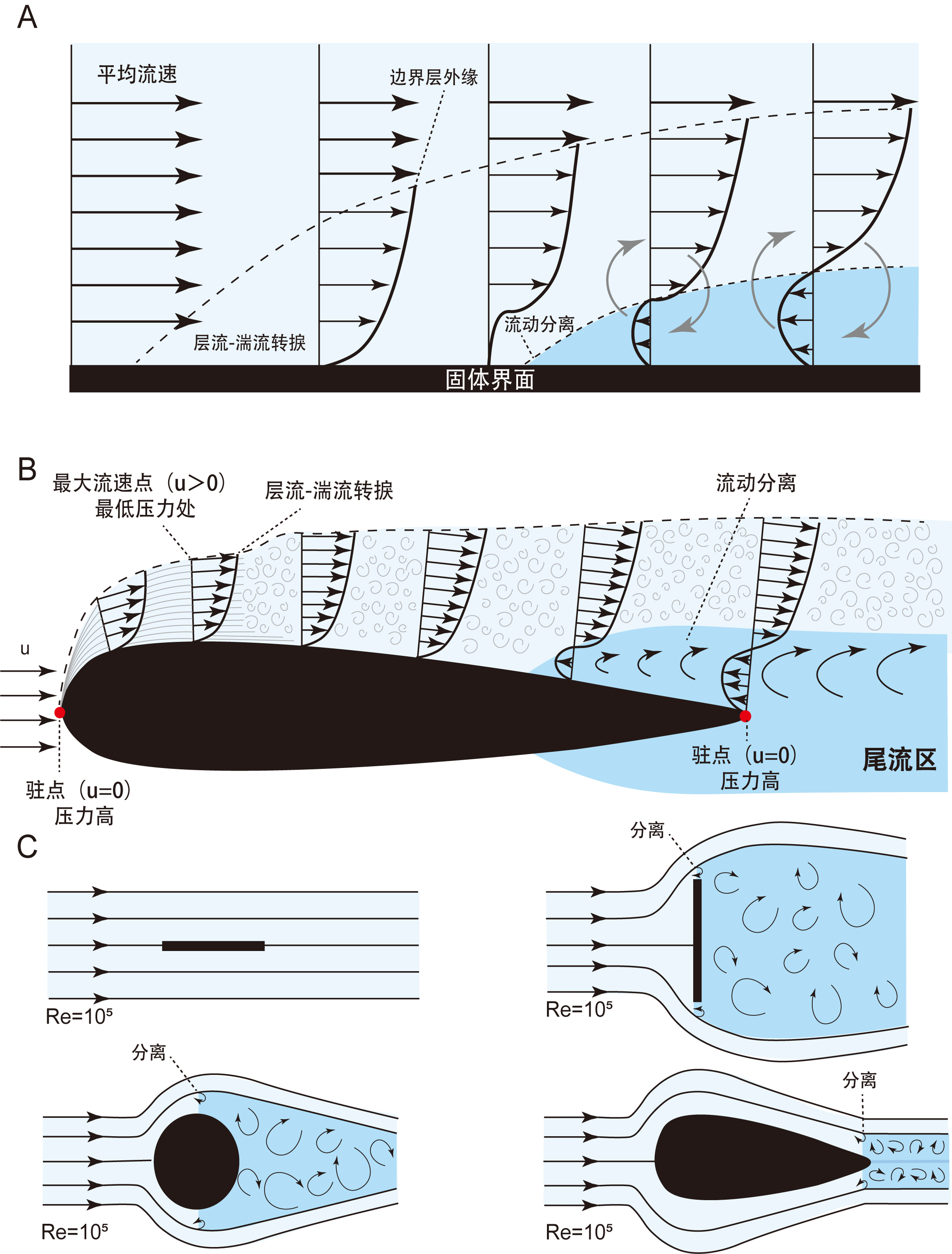
图3 计算流体动力学研究基本原理示意图。A,受逆压梯度作用的平板上边界层的发展阶段。箭头表示流动方向,长度表示水流流速,漩涡形成区或尾迹区为蓝色(修改自[19])。B,绕流线型物体的流动模式与边界层示意图。在物体前部,层流边界层的特征为平滑的流线。在层流-湍流转捩之后,湍流边界层的特征为不规则、涡旋状的流线。C,在相同雷诺数下(Re=105),绕不同物体的流动模式的理想化示意图,示边界层的发展与分离 [29]。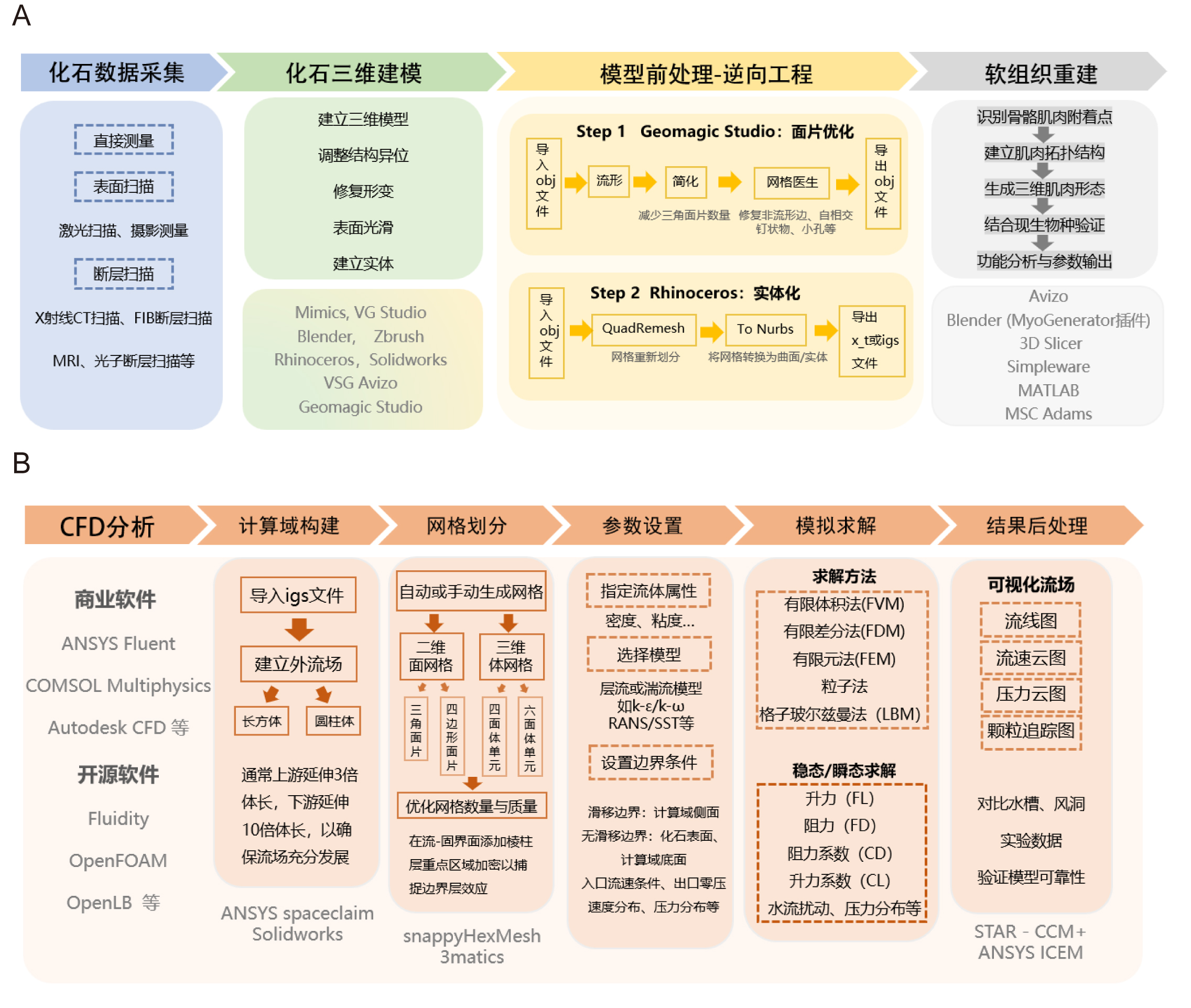
图4 古生物计算流体动力学研究流程图。A,化石三维数据获取与前处理流程。B,化石计算流体动力学分析过程与常用软件。
利益冲突
作者声明无利益冲突。
参考文献
- Coveney, P. V., Boon, J. P. and Succi, S. (2016). Bridging the gaps at the physics–chemistry–biology interface. Philos Trans A Math Phys Eng. Sci. 374(2080): 20160335. https://doi.org/10.1098/rsta.2016.0335
- Hoi, Y., Meng, H., Woodward, S. H., Bendok, B. R., Hanel, R. A., Guterman, L. R. and Hopkins, L. N. (2004). Effects of arterial geometry on aneurysm growth: three-dimensional computational fluid dynamics study. J Neurosurg. 101(4): 676–681. https://doi.org/10.3171/jns.2004.101.4.0676
- Anderson, J. D. and Wendt, J. (1995). Computational fluid dynamics (Vol. 206, p. 332). New York: McGraw-hill.
- Rahman, I. A. (2017). Computational fluid dynamics as a tool for testing functional and ecological hypotheses in fossil taxa. Palaeontology. 60(4): 451–459. https://doi.org/10.1111/pala.12295
- Shiino, Y., Kuwazuru, O. and Yoshikawa, N. (2009). Computational fluid dynamics simulations on a Devonian spiriferid Paraspirifer bownockeri (Brachiopoda): Generating mechanism of passive feeding flows. J Theor Biol. 259(1): 132–141. https://doi.org/10.1016/j.jtbi.2009.02.018
- Shiino, Y., Kuwazuru, O., Suzuki, Y., Ono, S. and Masuda, C. (2014). Pelagic or benthic? Mode of life of the remopleuridid trilobite Hypodicranotus striatulus. Bull Geosci. : 207–218. https://doi.org/10.3140/bull.geosci.1409
- Caromel, A. G., Schmidt, D. N., Phillips, J. C. and Rayfield, E. J. (2014). Hydrodynamic constraints on the evolution and ecology of planktic foraminifera. Mar Micropaleontol. 106: 69–78. https://doi.org/10.1016/j.marmicro.2014.01.002
- Rahman, I. A., Darroch, S. A. F., Racicot, R. A. and Laflamme, M. (2015). Suspension feeding in the enigmatic Ediacaran organism Tribrachidium demonstrates complexity of Neoproterozoic ecosystems. Sci Adv. 1(10): e1500800. https://doi.org/10.1126/sciadv.1500800
- Dynowski, J. F., Nebelsick, J. H., Klein, A. and Roth-Nebelsick, A. (2016). Computational Fluid Dynamics Analysis of the Fossil Crinoid Encrinus liliiformis (Echinodermata: Crinoidea). PLoS One. 11(5): e0156408. https://doi.org/10.1371/journal.pone.0156408
- Gutarra, S., Moon, B. C., Rahman, I. A., Palmer, C., Lautenschlager, S., Brimacombe, A. J. and Benton, M. J. (2019). Effects of body plan evolution on the hydrodynamic drag and energy requirements of swimming in ichthyosaurs. Proc R Soc Lond B Biol Sci. 286(1898): 20182786. https://doi.org/10.1098/rspb.2018.2786
- Liu, S., Smith, A. S., Gu, Y., Tan, J., Liu, C. K. and Turk, G. (2015). Computer Simulations Imply Forelimb-Dominated Underwater Flight in Plesiosaurs. PLoS Comput Biol. 11(12): e1004605. https://doi.org/10.1371/journal.pcbi.1004605
- BOTELLA, H. and FARIÑA, R. A. (2008). FLOW PATTERN AROUND THE RIGID CEPHALIC SHIELD OF THE DEVONIAN AGNATHANERRIVASPIS WAYNENSIS(PTERASPIDIFORMES: HETEROSTRACI). Palaeontology. 51(5): 1141–1150. https://doi.org/10.1111/j.1475-4983.2008.00801.x
- Ferrón, H. G., Martínez-Pérez, C., Rahman, I. A., Selles de Lucas, V., Botella, H. and Donoghue, P. C. (2020). Computational Fluid Dynamics Suggests Ecological Diversification among Stem-Gnathostomes. Curr Biol. 30(23): 4808–4813.e3. https://doi.org/10.1016/j.cub.2020.09.031
- Kogan, I., Pacholak, S., Licht, M., Schneider, J. W., Brücker, C. and Brandt, S. (2015). The invisible fish: hydrodynamic constraints for predator-prey interaction in fossil fishSaurichthyscompared to recent actinopterygians. Biol Open. 4(12): 1715–1726. https://doi.org/10.1242/bio.014720
- 曾军,张剑.(2025). 计算流体动力学在航空发动机研制中的应用. 燃气涡轮试验与研究, 38: 58—72.
- Khan, A. H., Ruiz Hussmann, K., Powalla, D., Hoerner, S., Kruusmaa, M. and Tuhtan, J. A. (2022). An open 3D CFD model for the investigation of flow environments experienced by freshwater fish. Ecol Inf. 69: 101652. https://doi.org/10.1016/j.ecoinf.2022.101652
- Falcucci, G., Amati, G., Fanelli, P., Krastev, V. K., Polverino, G., Porfiri, M. and Succi, S. (2021). Extreme flow simulations reveal skeletal adaptations of deep-sea sponges. Nature. 595(7868): 537–541. https://doi.org/10.1038/s41586-021-03658-1
- 冯文谦,李启剑,那琳,张景新,李越.(2024). 浅析大涡模拟在古生态学中的应用. 微体古生物学报, 41: 65–77.
- Fletcher, T., Altringham, J., Peakall, J., Wignall, P. and Dorrell, R. (2014). Hydrodynamics of fossil fishes. Proc R Soc Lond B Biol Sci. 281(1788): 20140703. https://doi.org/10.1098/rspb.2014.0703
- Fish, F. and Lauder, G. (2006). PASSIVE AND ACTIVE FLOW CONTROL BY SWIMMING FISHES AND MAMMALS. Annu Rev Fluid Mech. 38(1): 193–224. https://doi.org/10.1146/annurev.fluid.38.050304.092201
- Hao, J., Wang, J. and Lee, C. (2017). Assessment of vibration–dissociation coupling models for hypersonic nonequilibrium simulations. Aerosp Sci Technol. 67: 433–442. https://doi.org/10.1016/j.ast.2017.04.027
- Wang, J., Gao, Z., Lee, C. and Zhang, H. (2014). A decoupled procedure for convection-radiation simulation in scramjets. Sci China Technol Sci. 57(12): 2551–2566. https://doi.org/10.1007/s11431-014-5706-y
- Young, A., Gourlay, T., McKee, S. and Danton, M. H. (2014). Computational modelling of the hybrid procedure in hypoplastic left heart syndrome: A comparison of zero-dimensional and three-dimensional approach. Med Eng Phys. 36(11): 1549–1553. https://doi.org/10.1016/j.medengphy.2014.08.015
- Rahman, I. A. (2020). Computational Fluid Dynamics and its Applications in Echinoderm Palaeobiology. Cambridge University Press. Cambridge. 1–29. https://doi.org/10.1017/9781108893473
- Chamberlain, J. A. and Westermann, G. E. G. (1976). Hydrodynamic properties of cephalopod shell ornament. Paleobiology 2(4): 316–331. https://doi.org/10.1017/s0094837300004954
- Botella, H., Fariña, R. A. and Huera-Huarte, F. (2024). Delta wing design in earliest nektonic vertebrates. Commun Biol. 7(1): 1153. https://doi.org/10.1038/s42003-024-06837-8
- Dec, M. (2019). Hydrodynamic performance of psammosteids: new insights from computational fluid dynamics simulations. Acta Palaeontol Pol. 64: e2019. https://doi.org/10.4202/app.00623.2019
- Gai, Z., Li, Q., Ferrón, H. G., Keating, J. N., Wang, J., Donoghue, P. C. J. and Zhu, M. (2022). Galeaspid anatomy and the origin of vertebrate paired appendages. Nature. 609(7929): 959–963. https://doi.org/10.1038/s41586-022-04897-6
- Gutarra, S. and Rahman, I. A. (2021). The locomotion of extinct secondarily aquatic tetrapods. Biol Rev. 97(1): 67–98. https://doi.org/10.1111/brv.12790
- Troelsen, P. V., Wilkinson, D. M., Seddighi, M., Allanson, D. R. and Falkingham, P. L. (2019). Functional morphology and hydrodynamics of plesiosaur necks: does size matter?. J Vertebr Paleontol. 39(2): e1594850. https://doi.org/10.1080/02724634.2019.1594850
- Muscutt, L. E., Dyke, G., Weymouth, G. D., Naish, D., Palmer, C. and Ganapathisubramani, B. (2017). The four-flipper swimming method of plesiosaurs enabled efficient and effective locomotion. Proc R Soc Lond B Biol Sci. 284(1861): 20170951. https://doi.org/10.1098/rspb.2017.0951
- Bourke, J. M., Ruger Porter, W., Ridgely, R. C., Lyson, T. R., Schachner, E. R., Bell, P. R. and Witmer, L. M. (2014). Breathing Life Into Dinosaurs: Tackling Challenges of Soft‐Tissue Restoration and Nasal Airflow in Extinct Species. Anat Rec. 297(11): 2148–2186. https://doi.org/10.1002/ar.23046
- 王乐姣,郭颖,安晓晨,赵艳,王京盈. (2019). 孔子鸟三维复原与飞行性能数值模拟初探. 自然科学. 07: 590–595.
- Breder, C. (1926). The locomotion of fishes. Zoologica; Scientific Contributions of the New York Zoological Society. 4(5): 159–297.
- Webb, P. W. (1984). Form and Function in Fish Swimming. Sci Am. 251(1): 72–82. https://doi.org/10.1038/scientificamerican0784-72
- Sfakiotakis, M., Lane, D. and Davies, J. (1999). Review of fish swimming modes for aquatic locomotion. IEEE J Oceanic Eng. 24(2): 237–252. https://doi.org/10.1109/48.757275
- Webb, P. W. (1973). Kinematics of Pectoral Fin Propulsion in Cymatogaster Aggregata. J Exp Biol. 59(3): 697–710. https://doi.org/10.1242/jeb.59.3.697
- 魏倩芸. (2020). 基于CFD的胸/尾鳍协同推进仿生机器鱼水动力学特性分析. 兰州交通大学.
- 魏倩芸,李宗刚,高溥,杜亚江. (2021). 仿生机器鱼胸尾鳍协同推进的水动力学分析. 船舶工程. 43: 43–49.
- Borazjani, I. and Sotiropoulos, F. (2010). On the role of form and kinematics on the hydrodynamics of self-propelled body/caudal fin swimming. J Exp Biol. 213(1): 89–107. https://doi.org/10.1242/jeb.030932
- Gibson, B. M., Rahman, I. A., Maloney, K. M., Racicot, R. A., Mocke, H., Laflamme, M. and Darroch, S. A. F. (2019). Gregarious suspension feeding in a modular Ediacaran organism. Sci Adv. 5(6): eaaw0260. https://doi.org/10.1126/sciadv.aaw0260
- Ko, H., Girma, A., Zhang, Y., Pan, Y., Lauder, G. and Nagpal, R. (2025). Beyond planar: fish schools adopt ladder formations in 3D. Sci Rep. 15(1): e1038/s41598–025–06150–2. https://doi.org/10.1038/s41598-025-06150-2
- Mao, X., Wang, J., Mao, X. and Deng, J. (2025). A hybrid numerical model for the collective motion of fish groups. J Fluid Mech. 1009: e216. https://doi.org/10.1017/jfm.2025.216
- Marcé-Nogué, J. (2022). One step further in biomechanical models in palaeontology: a nonlinear finite element analysis review. PeerJ. 10: e13890. https://doi.org/10.7717/peerj.13890
- Lautenschlager, S. (2020). Multibody dynamics analysis (MDA) as a numerical modelling tool to reconstruct the function and palaeobiology of extinct organisms. Palaeontology. 63(5): 703–715. https://doi.org/10.1111/pala.12501
- Hutchinson, J. R. and Garcia, M. (2002). Tyrannosaurus was not a fast runner. Nature. 415(6875): 1018–1021. https://doi.org/10.1038/4151018a
- Lautenschlager, S. (2016). DIGITAL RECONSTRUCTION OF SOFT-TISSUE STRUCTURES IN FOSSILS. The Paleontological Society Papers 22: 101–117. https://doi.org/10.1017/scs.2017.10
- Rigby, S. and Tabor, G. (2006). The use of computational fluid dynamics in reconstructing the hydrodynamic properties of graptolites. GFF 128(2): 189–194. https://doi.org/10.1080/11035890601282189
您对这篇实验方法有问题吗?
在此处发布您的问题,我们将邀请本文作者来回答。同时,我们会将您的问题发布到Bio-protocol Exchange,以便寻求社区成员的帮助。